Контрольная работа: Вопросы экологии применительно к лесному хозяйству
Раздел: Рефераты по экологии
Тип: контрольная работа
ЭКОЛОГИЯ
Контрольная работа № 1
Вопрос № 8.
Вода, как среда обитания животных организмов: плотность, давление, кислород, освещенность, солевой режим, течения, температура. Отличия от воздушной среды. Приспособления растений и животных.
Благодаря широкой распространённости воды и её роли в жизни людей, она издавна считалась первоисточником жизни. Представление философов античности о воде как о начале всех вещей нашло отражение в учении Аристотеля (4 в. до н. э.) о четырёх стихиях (огне, воздухе, земле и воде), причём вода считалась носителем холода и влажности. Вплоть до конца 18 века в науке существовало представление о воде, как об индивидуальном химическом элементе. В 1781—82 годах английский учёный Г. Кавендиш впервые синтезировал воду, взрывая электрической искрой смесь водорода и кислорода, а в 1783 году французский учёный А. Лавуазье повторив эти опыты, впервые сделал правильный вывод, что вода есть соединение водорода и кислорода. В 1785 году Лавуазье совместно с французским учёным Ж. Менье определил количественный состав воды. В 1800 году английские учёные У. Николсон и А. Карлейль разложили воду на элементы электрическим током. Таким образом, анализ и синтез воды показали сложность её состава и позволили установить для неё формулу H2O. Изучение физических свойств воды началось ещё до установления её состава в тесной связи с другими научно-техническими проблемами. В 1612 году итальянский учёный Г. Галилей обратил внимание на меньшую плотность льда сравнительно с жидкой водой как на причину плавучести льда. В 1665 году голландский учёный Х. Гюйгенс предложил принять температуру кипения и температуру плавления воды за опорные точки шкалы термометра. В 1772 году французский физик Делюк нашёл, что максимум плотности воды лежит при 4°С; при установлении в конце 18 века метрической системы мер и весов это наблюдение было использовано для определения единицы массы – килограмма. В связи с изобретением паровой машины французские учёные Д. Араго и П. Дюлонг (1830 г.) изучили зависимость давления насыщенного пара воды от температуры. В 1891—97 годах Д. И. Менделеев дал формулы зависимости плотности воды от температуры. В 1910 году американский учёный П. Бриджмен и немецкий учёный Г. Тамман обнаружили у льда при высоком давлении несколько полиморфных модификаций. В 1932 году американские учёные Э. Уошберн и Г. Юри открыли тяжёлую воду. Развитие физических методов исследования позволило существенно продвинуться в изучении структуры молекул воды, а также строения кристаллов льда. В последние десятилетия особое внимание учёных привлекает структура жидкой воды и водных растворов.
Физические свойства и строение воды. Важнейшие физические константы воды приведены в таблице 1. Тройная точка для воды, где находятся в равновесии жидкая вода, лёд и пар, лежит при температуре +0,01°С и давлении 6,03·10-3 атм.
Многие физические свойства воды обнаруживают существенные аномалии. Как известно, свойства однотипных химических соединений у элементов, находящихся в одной и той же группе периодической системы Менделеева, изменяются закономерно. В ряду водородных соединений элементов VI группы (H2Te, H2Se, H2S, H2O) температуры плавления и кипения закономерно уменьшаются лишь у первых трёх; для воды эти температуры аномально высоки. Плотность воды в интервале 100—4°С нормально возрастает, как и у огромного большинства других жидкостей. Однако, достигнув максимального значения 1,0000 г/см3 при +3,98°С, при дальнейшем охлаждении уменьшается, а при замерзании скачкообразно падает, тогда как почти у всех остальных веществ кристаллизация сопровождается увеличением плотности. Вода способна к значительному переохлаждению, т. е. может оставаться в жидком состоянии ниже температуры плавления (даже при —30°С). Удельная теплоёмкость, удельная теплота плавления и кипения воды аномально высоки по сравнению с другими веществами, причём удельная теплоёмкость воды минимальна при 40°С. Вязкость воды с ростом давления уменьшается, а не повышается, как следовало бы ожидать по аналогии с другими жидкостями. Сжимаемость воды крайне невелика, причём с ростом температуры уменьшается.
Таблица 1. Физические свойства воды
| Свойство | Значение Воздух для ср. |
|
Плотность, г/см3 |
|
| лёд | 0,9168 (0°С) |
| жидкость |
0,99987 (0°С) 1,0000 (3,98°С) 0,99823 (20°С) 0,001248 (20°С) |
| Пар насыщенный |
0,5977 кг/м3 (100°С) |
| Температура плавления | 0°С |
| Температура кипения | 100°С |
| Критическая температура | 374,15°С —140,7°С |
| Критическое давление |
218,53 кгс/см2 3,7 Мн/м2 (37,2 am) |
| Критическая плотность |
0,325 г/см3 |
| Теплота плавления | 79,7 кал/г |
| Теплота испарения | 539 кал/г (100°С) |
|
Удельная теплопроводность, кал/(см·сек·град) |
|
| лёд |
5,6·10-3 (0°С) |
| жидкость |
1,43·10-3 (0°С) 1,54·10-3 (45°С) |
| Пар насыщенный |
5,51·10-5 (100°С) |
|
Уд. электропроводность, ом--1·см-1 |
|
| лёд |
0,4·10-8 (0°С) |
| жидкость |
1,47·10-8 (0°С) 4,41·10-8 (18°С) 18,9·10-8 (50°С) |
|
Удельная теплоёмкость Кал/(г·град) |
|
| жидкость |
1,00 (15°С) 10,045·103 дж/(кг·К) |
| Пар насыщенный | 0,487 (100°С) |
| Диэлектрическая проницаемость | |
| лёд | 74,6 (°С) |
| жидкость | 81,0 (20°С) 1,000059 (0°С). |
| Пар насыщенный | 1,007 (145°С) |
| Вязкость, спз | |
| Жидкость |
1,7921 (0°С) 0,000171 (0°С) 0,284 (100°С) |
|
Поверхностное натяжение жидкой воды на границе с воздухом, дин/см |
74,64 (0°С) 62,61 (80°С) |
|
Показатель преломления (D — линия |
1,33299 (20°С) 1,00029 |
Примечание: 1 кал/(см·сек·град) = 418,68 вт/(м·К); 1 ом-–1·см-–1 = 100 сим/м;
1 кал/(г·град) =.4,186 кдж (кг·К); 1 спз = 10—3н·сек/м2; 1 дин/см = 10–3н/м.
Аномалии физических свойств воды связаны со структурой её молекулы и особенностями межмолекулярных взаимодействий в жидкой воде и льде. Три ядра в молекуле В. образуют равнобедренный треугольник с протонами в основании и кислородом в вершине (Рис. 1, а). Распределение электронной плотности в молекуле В. таково (Рис 1, б, в), что создаются 4 полюса зарядов: 2 положительных, связанных с атомами водорода, и 2 отрицательных, связанных с электронными облаками необобществлённых пар электронов атома кислорода. Указанные 4 полюса зарядов располагаются в вершинах тетраэдра (Рис 1, г). Благодаря этой полярности вода имеет высокий дипольный момент(1,86 D), а четыре полюса зарядов позволяют каждой молекуле воды образовать четыре водородные связи с соседними (такими же) молекулами (например, в кристаллах льда).м
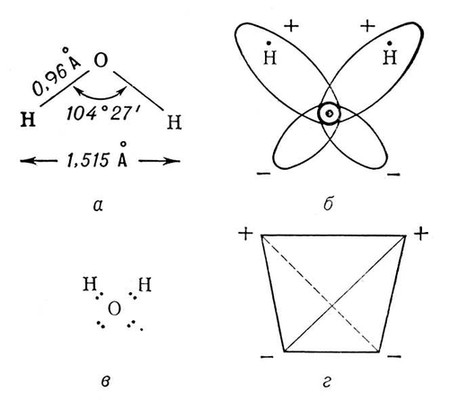
Рис. 1. Структура молекулы воды: а — геометрия молекулы H2O (в парообразном состоянии); б — электронные орбиты в молекуле H2O; в — электронная формула молекулы H2O (видны необобществленные электронные пары); г — четыре полюса зарядов в молекуле H2O расположены в вершинах тетраэдра.
Вода входит в состав всех живых организмов, причём в целом в них содержится лишь вдвое меньше воды, чем во всех реках Земли. В живых организмах количество воды, за исключением семян и спор, колеблется между 60 и 99,7% по массе. По словам французского биолога Э. Дюбуа-Реймона, живой организм есть l'eau animée (одушевлённая вода). Все воды Земли постоянно взаимодействуют между собой, а также с атмосферой, литосферой и биосферой.
Вода в природных условиях всегда содержит растворённые соли, газы и органические вещества. Их количественный состав меняется в зависимости от происхождения воды и окружающих условий. При концентрации солей до 1 г/кг воду считают пресной, до 25 г/кг — солоноватой, свыше — солёной.
Наименее минерализованными водами являются атмосферные осадки (в среднем около 10—20 мг/кг), затем пресные озёра и реки (50—1000 мг/кг). Солёность океана колеблется около 35 г/кг; моря имеют меньшую минерализацию (Чёрное 17—22 г/кг; Балтийское 8—16 г/кг; Каспийское 11—13 г/кг). Минерализация подземных вод вблизи поверхности в условиях избыточного увлажнения составляет до 1 г/кг, в засушливых условиях до 100 г/кг, в глубинных артезианских вода минерализация колеблется в широких пределах. Максимальные концентрации солей наблюдаются в соляных озёрах (до 300 г/кг) и глубокозалегающих подземных водах. (до 600 г/кг).
В пресных водах обычно преобладают ионы HCO3-, Са2+ и Mg2+. По мере увеличения общей минерализации растет концентрация ионов SO42-, Cl-, Na+ и К+. В высо-коминерализованных водах преобладают ионы Cl- и Na+, реже Mg2+ и очень редко Ca2+. Прочие элементы содержатся в очень малых количествах, хотя почти все естественные элементы периодической системы найдены в природных водах.
Из растворённых газов в природных водах присутствуют азот, кислород, двуокись углерода, благородные газы, редко сероводород и углеводороды. Концентрация органических веществ невелика — в среднем в реках около 20 мг/л, в подземных водах ещё меньше, в океане около 4 мг/л. Исключение составляют воды болотные и нефтяных месторождений и воды, загрязнённые промышленными и бытовыми стоками, где количество их бывает выше. Качественный состав органических веществ чрезвычайно разнообразен и включает различные продукты жизнедеятельности организмов, населяющих воду, и соединения, образующиеся при распаде их остатков.
Первоисточниками солей природных вод являются вещества, образующиеся при химическом выветривании изверженных пород (Ca2+, Mg2+, Na+, К+ и др.), и вещества, выделявшиеся на протяжении всей истории Земли из её недр (CO2, SO2, HCI, NH3 и др.). От разнообразия состава этих веществ и условий, в которых происходило их взаимодействие с водой, зависит состав воды. Громадное значение для состава воды имеет и воздействие живых организмов.
В связи с существованием двух стабильных изотопов у водорода (1H и 2H, обычно обозначаемые Н и D) и трёх у кислорода (16O,17O и 18O) известно 9 изотопных разновидностей воды, которые находятся в природной воде в среднем в следующих соотношениях (в молярных %): 99,73 H216O; 0,04 H217O; 0,20 H218O, 0,03 HD’16O, а также 10-5—10-15%(суммарно) HD17O, HD18O, D216O, D217O, D218O. Особый интерес представляет тяжелая вода D2O, содержащая дейтерий. В водах Земли находится всего13—20 кг «сверхтяжёлой» воды. содержащей радиоактивный изотоп водорода — тритий (3H, или Т).
Вода в организме — основная среда (внутриклеточная и внеклеточная), в которой протекает обмен веществ у всех растений, животных и микроорганизмов, а также субстрат ряда химических ферментативных реакций. В процессе фотосинтеза вода вместе с углекислым газом вовлекается в образование органических веществ и, таким образом, служит материалом для создания живой материи на Земле.
Таблица 2. Содержание воды в различных организмах, их органах и тканях
| Организмы, органы, ткани | Содержание воды, % |
| Растения (наземные) | |
| верхушка растущего побега | 91—93 |
| листья . . . . . . . . . . . . . . . . . | 75—86 |
| Семена злаков . . . . . . . . . . . | 12—14 |
| Водоросли . . . . . . . . . . . . . . . | 90—98 |
| Мхи, лишайники . . . . . . . . . . | 5—7 |
| Медузы . . . . . . . . . . . . . . . . . | 95–98 |
| Дождевые черви . . . . . . . . . . | 84 |
| Насекомые | |
| взрослые . . . . . . . . . . . . . . . | 45—65 |
| личинки . . . . . . . . . . . . . . . . | 58—90 |
| Рыбы . . . . . . . . . . . . . . . . . . . | 70 |
|
Млекопитающие (в т. ч. человек) |
63—68 |
| скелет | 20—40 |
| мышцы | 75 |
| печень | 75 |
| Мозг человека | |
| серое вещество | 84 |
| белое вещество | 72 |
Вода обеспечивает тургор тканей, перенос питательных веществ и продуктов обмена (кровь, лимфа, сок растений), физическую терморегуляцию и другие процессы жизнедеятельности. Жизнь, вероятно, возникла в водной среде. В ходе эволюции различные водные животные и водные растения вышли на сушу и приспособились к наземному образу жизни; тем не менее и для них вода — важнейший компонент внешней среды. Жизнь без воды невозможна. При недостатке воды жизнедеятельность организмов нарушается. Лишь покоящиеся формы жизни — споры, семена — хорошо переносят длительное обезвоживание. Растения при отсутствии воды увядают и могут погибнуть, но чувствительность различных растений к недостатку воды неодинакова. Животные, если лишить их воды, быстро погибают: упитанная собака может прожить без пищи до 100 дней, а без воды — менее 10. Содержание воды в организмах велико (см. таблицу 2).
В жидкостях организма — межклеточных пространствах, лимфе, крови, пищеварительных соках, соке растений и др. — содержится свободная вода. В тканях животных и растений вода находится в связанном состоянии — она не вытекает при рассечении органа. Вода способна вызывать набухание коллоидов, связываться с белком и другими органическими соединениями, а также с ионами, входящими в состав клеток и тканей (гидратационная вода). Молекулы воды, находящиеся внутри клеток, но не входящие в состав гидратационных оболочек ионов и молекул, представляют иммобильную воду, легче гидратационной вовлекаемую в общий круговорот воды в организме.
Вода физиологически необходима любой протоплазме и с экологической точки зрения является лимитирующим фактором, как в наземных, так и в водных местообитаниях, если там ее количество подвержено резким изменениям (приливы, отливы) или происходит ее потеря организмом в сильно соленой воде осмотическим путем.
В наземно-воздушной среде этот абиотический фактор характеризуется величиной количества осадков, влажности, иссушающими свойствами воздуха и доступной площадью водного запаса.
Количество атмосферных осадков обусловлено физико-географическими условиями и неравномерно распределено на земном шаре. Но для организмов важнейшим лимитирующим фактором является распределение осадков по сезонам года. В умеренных широтах даже при достаточном количестве годовых осадков их неравномерное распределение может привести к гибели растений от засухи (лесных пожаров) или, наоборот, от переувлажнения. В тропической зоне организмам приходится переживать влажные и сухие сезоны, регулирующие их сезонную активность при постоянной почти круглый год температуре.
Адаптированные к условиям пустыни растения содержат ингибитор прорастания, который вымывается лишь при определенном количестве осадков, достаточном для вегетации (например, 10 мм) и тогда только прорастает. Начинается кратковременное «цветение пустыни» (обычно весной).
Иссушающее действие воздуха наиболее важное экологическое значение имеет для растений. Подавляющее большинство растений всасывает воду корневой системой из почвы. Иссушение почвы затрудняет всасывание. Адаптация растений к этим условиям – увеличение всасывающей силы и активной поверхности корней. Величина этой силы у корней умеренной зоны от 2 до 4 ∙ 106 Па, а у растений сухих областей – до 6 ∙ 106 Па. Как только выбрана вода в данном объеме, корни растут далее в глубь и стороны и корневая система может достигнуть, например, у злаков длины 13 км на 1000 см3 почвы (без корневых волосков).
Вода расходуется на фотосинтез, всего около 0,5 % всасывается клетками, а 97 – 99 % ее уходит на транспирацию – испарение через листья. При достатке воды и питательных веществ рост растений пропорционален транспирации, а ее эффективность будет наивысшей. Эффективность транспирации – это отношение прироста вещества (чистой продукции) к количеству транспирированной воды. Измеряется она в граммах сухого вещества на 1000 см3 воды. Для большинства растений она равна двум, т. е. на получение каждого грамма живого вещества тратится 500 г воды, даже для большинства засухоустойчивых. Основная форма адаптации – не снижение транспирации, а прекращение роста в период засухи.
В нижних ярусах тропических дождевых лесов, где 100%-ная относительная влажность, есть растения с приспособлениями для потери воды, а в пустынях у некоторых растений водный баланс не нарушается даже в период непродолжительной засухи, и т. д. В зависимости от способов адаптации растений к влажности выделяют несколько экологических групп, например: гигрофиты – наземные растения, живущие в очень влажных почвах и в условиях повышенной влажности (рис, папирус); мезофиты – переносят незначительную засуху (древесные растения различных климатических зон, травянистые растения дубрав, большинство культурных растений и др.); ксерофиты – растения сухих степей и пустынь, способные накапливать влагу в мясистых листьях и стеблях – суккуленты (алоэ, кактусы и др.), а также обладающие большой всасывающей силой корней и способные снижать транспирацию с узкими мелкими листьями – склерофиты.
Среди суккулентов наблюдается явление конвергенции – растения, относящиеся к разным видам, имеют практически одинаковую форму: африканский молочай и кактус имеют шарообразную форму, обеспечивающую наименьшую поверхность испарения.
Водная среда в среднем в 800 раз плотнее воздуха; этим объясняется возможность существования в воде животных, прозрачное, студенистое тело которых лишено прочных покровов или поддерживающего скелетного аппарата (медузы, сифонофоры, гребневики, сальпы и др.). Плотностью воды обусловлены и характерные для многих водных животных способы движения посредством ресничек или жгутиков (у большинства простейших, некоторых червей, кишечно-полостных и др., а также у личиночных форм губок, кишечно-полостных, червей, моллюсков, иглокожих и др.). Большая плотность воды позволяет очень мелким водным животным (планктон), способным лишь к слабым активным движениям, держаться в толще воды при помощи несложных приспособлений в виде крошечных пузырьков воздуха или капелек жира в их теле, или длинных, тонких выростов, увеличивающих поверхность тела. Только среди водных животных встречаются неподвижные прикреплённые формы, что обусловлено подвижностью воды и, следовательно, постоянным приносом находящейся в ней пищи в виде живых и отмерших планктонных организмов, так же как и разносом оплодотворённых яиц и личинок, который обеспечивает расселение прикрепленных форм.
Водные растения, также как и животные, обитающие в воде, в ходе эволюции своеобразно приспособились к данной среде обитания. Среди них различают гидрофиты — растения, погружённые в воду только нижней частью, и гидатофиты — растения, полностью или большей своей частью погруженные в воду. Обитание в водной среде обусловило особые черты организации водных растений: значительное увеличение поверхности тела в сравнении с его массой, что облегчает поглощение необходимых количеств кислорода и других газов, которых в воде содержится меньше, чем в воздухе. Увеличение поверхности тела достигается развитием больших тонких листьев (некоторые рдесты), расчленением листовой пластинки на тонкие нитевидные участки (уруть, роголистники, водные лютики); продырявливанием листьев или сильным развитием воздухоносных полостей и больших межклетников. У водных растений сильно развита разнолистность (гетерофилия): подводные, плавающие и воздушные листья на одном и том же растении значительно различаются как по внутреннему, так и по внешнему строению. Так, подводные листья не имеют устьиц; у плавающих на поверхности воды листьев устьица находятся только на верхней стороне, у воздушных листьев устьица — на обеих сторонах. Большая плотность водной среды обусловливает слабое развитие механических элементов в листьях и стеблях водных растений; немногочисленные механические элементы, имеющиеся в стеблях, расположены ближе к центру, что придаёт им большую гибкость. Так как интенсивность света в воде резко снижается, у многих водных растений в клетках эпидермиса имеются хлорофилловые зёрна. У водных растений слабо развиты или даже отсутствуют сосуды в проводящих пучках. Слабо развита и корневая система, а корневые волоски отсутствуют. Почти все водные растения — многолетники, размножающиеся вегетативно. Некоторые водные растения (наяда, роголистник) опыляются под водой; у других цветки поднимаются над водой, где и происходит опыление. Некоторые водные растения приспособились к периодическому высыханию водоёмов (например, частуха, стрелолист, жеруха).
Вопрос № 18
Динамика ценопопуляций растений: монокарпики и поликарпики, инвазия, запас диаспор в почве, семенная продуктивность, популяционные волны, самоизреживание, сукцессии, флюктуации, гомеостаз.
Ценопопуляция по определению – это составная часть биогеоценоза. Еще в XVII веке было установлено, что численность популяций растет по закону геометрической прогрессии, а уже в конце XVIII века Томас Мальтус (1766 – 1834 гг.) выдвинул свою известную теорию о росте народонаселения в геометрической прогрессии. Эта закономерность выражается кривой, изображенной на рис. 2.
На современном математическом языке эта кривая отражает экспоненциальный рост численности организмов и описывается уравнением:
Nt = N0 ert ,
где Nt – численность популяции в момент времени t;
N0 – численность популяции в начальный момент времени t 0;
e – основание натурального логарифма (2,7182);
r – показатель, характеризующий темп размножения особей в данной популяции.
Экспоненциальный рост возможен только тогда, когда r имеет постоянное численное значение, так как скорость роста популяции пропорциональна самой численности:
ΔN / Δ t = r N, где r = const.
Если численность отложить в логарифмическом масштабе, то кривая приобретает вид прямой линии (рис. 2, b).
Таким образом, экспоненциальный рост численности популяции – это рост численности ее особей в не изменяющихся условиях.
Условия, сохраняющиеся длительное время постоянными, невозможны в природе. Если бы это было не так, то, например, обычные бактерии могли бы дать такую массу органического вещества, которая могла покрыть весь земной шар слоем толщиной в 2 метра за 2 часа.
Однако такого в природе не происходит, так как существует множество ограничивающих факторов. Но есть примеры, когда при замедлении роста, т. е. при снижении r, экспоненциальный рост сохраняется, может он возникать и на коротких отрезках жизни популяций.
Чтобы иметь полную картину динамики численности популяции, а так же рассчитать скорость ее роста, необходимо знать величину так называемой чистой скорости воспроизводства (R0), которая показывает, во сколько раз увеличивается численность популяции за одно поколение, за время его жизни T.
R0 = Nr / N0 ,
где Nr – численность нового поколения;
N0 – численность особей предшествующего поколения;
R0 – чистая скорость воспроизводства, показывающая также, сколько вновь родившихся особей приходится на одну особь поколения родителей. Если R0 = 1, то популяция стационарная, численность ее сохраняется постоянной.
Скорость роста популяции обратно пропорциональна длительности жизни поколения:
r = ln R0 / T,
отсюда ясно, что чем раньше происходит размножение организмов, тем больше скорость роста популяции. Беря во внимание, например, этологическую структуру популяций монокарпических и поликарпических растений, выражающую характер их организации, можно заметить тот факт, что скорость роста популяции монокарпиков значительно ниже скорости роста популяции поликарпиков. Это объясняется тем, что
R0 м << R0 п ,
где R0 м – чистая скорость воспроизводства монокарпических растений;
R0 п – чистая скорость воспроизводства поликарпических растений.
Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (r = 0), либо ее уменьшить. Например, к уменьшению численности популяции может привести воздействие инвазионных заболеваний, вызванных заражением растений паразитами животной природы. Инвазия может произойти активно, когда паразит нападает или внедряется в организм так называемого хозяина через поврежденные и неповрежденные ткани, или пассивно, когда паразит вносится в организм с водой, воздухом, почвенными растворами. Основными источниками заражения являются сточные воды предприятий пищевой и кожевенной промышленности, бытовые и промышленные свалки, кладбища, канализационная сеть, поля орошения и др. Из этих источников разнообразные органические соединения и патогенные микроорганизмы попадают в почву, горные породы и подземные воды. По данным санэпидстанций, патогенные кишечные палочки обнаруживаются в подземных водах на глубине до 300 метров от поверхности земли.
Состав и концентрация почвенного раствора определяют реакцию среды, показателем которой является величина pH. Наиболее благоприятной для растений и почвенных животных является нейтральная среда (pH = 7).
Однако в уменьшении численности популяций обычно играют роль вполне естественные ограничивающие или лимитирующие факторы (закон минимума Ю. Либиха, закон независимости факторов В. Р. Вильямса и закон толерантности В Шелфорда). В результате экспоненциальный рост замедляется или останавливается полностью и J – образная кривая экспоненциального роста как бы останавливается и выполаживается, превращаясь в так называемую S – образную кривую (рис. 3).
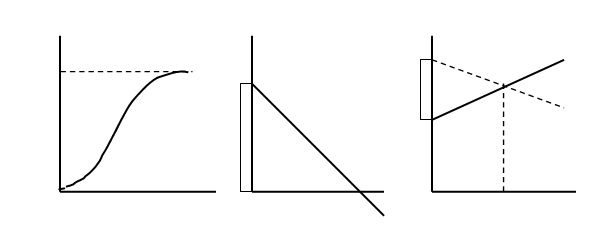
![]()
а r б b, d в
d
K
rmax
b
r max
N0 N N
0 Время, t 0 K 0 K
Рис. 3. Логистическая модель роста популяции: а – кривая роста численности (N); б – зависимость удельной скорости роста (r) от численности (N); в – зависимость рождаемости (b) и смертности (d) от численности; К – предельная численность.
В природе так и происходит: экспоненциальный рост наблюдается какое-то достаточно короткое время, после чего ограничивающие факторы его стабилизируют, и дальнейшее развитие популяции идет по логистической модели, что и описывается S – образной, или логистической кривой роста популяции.
В основе логистической модели (рис. 3) лежит простое допущение, что скорость роста популяции (ra) линейно снижается по мере роста численности вплоть до нуля при некой численности К. Итак, при начальной численности Na (близкой к нулю) скорость роста имеет максимальное значение r max , а при N = K, r a = 0. В результате решения уравнения логистической кривой получается зависимость:
Nt = K / (1 + e α – r max ∙ t ),
где Nt – численность популяции в момент времени t;
α – постоянная интегрирования.
Величины К называют еще емкостью среды в отношении особей данной популяции. Здесь речь идет о биологической емкости среды – степени способности природного или природно-антропогенного окружения обеспечить нормальную жизнедеятельность (дыхание, питание, размножение, отдых и т.д.) определенному числу организмов и их сообществ без заметного нарушения самого окружения.
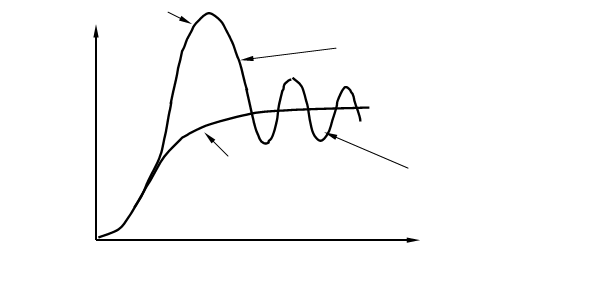
Однако плато на S – образной кривой далеко не всегда бывает гладким, потому что колебания численности происходят постоянно, что отражается в виде колебаний кривой вокруг асимптоты К (рис. 4), эти колебания называются популяционными волнами или флюктуациями численности. Данный термин был введён русским биологом С. С. Четвериковым в 1915 г. Подобные колебания численности могут быть сезонными или несезонными, повторяющимися через различные промежутки времени; обычно они тем длиннее, чем продолжительнее цикл развития организмов. Часто волны жизни сопровождаются колебаниями ареала популяций. Четвериков указал на эволюционное значение волн жизни в качестве фактора, могущего изменять направление и интенсивность отбора, а также концентрации генов, содержащихся в популяциях. Популяционные волны (один из 4 элементарных эволюционных факторов — мутационный процесс, популяционные волны, изоляция и естественный отбор). Основное значение популяционных волн сводится к случайным изменениям концентраций (особенно невысоких) различных мутаций и генотипов, содержащихся в популяциях, а также к ослаблению давления отбора при увеличении и его усилению при уменьшении численности особей в популяции. Под термином «популяционные волны» иногда (советский геолог Б. Л. Личков и некоторые другие ученые) также подразумевают этапы развития растительного и животного мира, примерно соответствующие смене геологических циклов.
Повышение популяцией
потенциальной емкости
«Крах» популяции
или вымирание
Число
особей
Потенциальная Популяционные
емкость волны
Время
Рис. 4. Преобразование J –образной кривой роста численности популяции в S – образную кривую при ограничивающем воздействии лимитирующих факторов.
Главенствующее значение для естественного возобновления ценопопуляции имеет запас диаспор в почве, наличие подроста и занос зачатков. В ненарушенной лесной среде практически исключена смена лесного типа растительности на луговой, степной или болотный. Если же ширина окна превышает две и более высоты сомкнутого окружающего полога, то возникает нелесная среда, где преимущество заселения получают вначале травяной и моховой типы растительности, и только потом заселяются пионерные береза, осина, иногда сосна, при этом частично или навсегда исчезают многие лесные виды растений, животных и, вероятно, микроорганизмов.
Можно наблюдать следующие закономерности возникновения возобновления сообществ. Почвенный запас семян хорошо защищен от поражения микроскопическими грибковыми организмами, неспорообразующими бактериями, спорообразующими бактериями, актиномицетами, но недостаточно – от представителей лесной фауны, особенно видов, питающихся в основном семенами. Случайным является попадание семян в оптимальные условия прорастания, совпадающие с генетической наследственностью. При прорастании семени проросток сразу вступает в тесные взаимоотношения с микробиоценозом, эдафотопом и климатопом, т.е. любое растение образует с биоценозом консорцию. В этот период наблюдается массовая гибель растений из-за несовпадения консорциативных и неблагоприятных экотопических связей. Объем пространства, занимаемый проростками и ювенильными древесными растениями, невелик, и конкуренции за свет и почву между ними нет. Ресурсы среды у них могут отнять мхи, травы и взрослые древесные особи. Это предопределяет всегда недостаточное число возобновления на начальном этапе. Даже количество возобновления, равное 2,5 млн. шт./га, означает 40 см2 (5 x 8 см), приходящихся на одно растение. Следовательно, энергию роста (бонитет) на начальном этапе полностью определяют почвенное богатство и сопряженность с микробоценозом. В это время закладывается жизненная стратегия роста и развития (так называемые классы Крафта – деление деревьев в лесу на пять классов по росту и развитию: прегосподствующие, господствующие, умеренно развитые, заглушенные и подпологовые) каждой особи. По мере роста особь увеличивает занимаемое ею пространство до начала смыкания кронами с другими особями, после чего начинается закономерная межвидовая и видовая ценопопуляционная конкуренция между особями в биогруппе вначале за свет надземного пространства, позже присоединяется конкуренция корневых систем особей за почвенные ресурсы. По отношению к каждой особи этот процесс получил название «давление среды», приводящее к изреживанию в биогруппах и в насаждении в целом. Выжившие особи увеличивают консорциативные связи, вступая в тесные взаимоотношения со всем биоценозом, формируя собственную свиту растений, животных, грибов, микроорганизмов и изменяя внутреннюю среду биогеоценоза в выгодном для себя отношении. В метрическом исчислении это выражается двумя показателями – высотой и занимаемым воздушным и почвенным объемами, т. е. числом особей на 1 га. Чем беднее почва, тем больше необходимый занимаемый особью объем почвы, в связи, с чем площадь крон отстает от площади корневой системы, в результате исключается стопроцентная сомкнутость крон, а значит не полностью создается лесная среда. Особенно это видно в переходных типах растительности. При любом возрасте конкурентные отношения выражаются высотой и числом стволов, что позволяет сравнивать с высокой степенью корреляции эти показатели при разных бонитетах и возрастах.
Все модификаторы стремятся в течение первоначальных 20 лет занять ведущее место в верхнем пологе. Если коренные породы сосна, дуб, ель осуществляют это за счет долголетия, то короткоживущие пионерные береза и осина – путем интенсификации природных ресурсов в первое пятилетие, увеличивая прирост в высоту на относительно богатых почвах на один класс бонитета, по сравнению с коренными, и одновременно формируя тонкий ствол и небольшой объем кроны и сучьев. Рост в высоту эдификаторов сдерживают следующие лимитирующие факторы:
· годовая теплообеспеченность в лесной зоне снижается с юга на север на 0,51 о С на 1о широты, что приводит к падению производительности на 0,2 – 0,3 класса бонитета на каждый градус широты на богатых почвах;
· при достижении 7 – 10 метров многие виды вступают в период плодоношения, затрачивая ресурсы на семяношение;
· по достижении 15 м и выше увеличение кроны приводит к необходимости наращивать толщину ствола во избежание ветролома;
· при высоте 20 метров и выше требуется увеличение корневой массы для избежания ветровала.
В зоне смешанных лесов Русской равнины и подзоне южной тайги рост в высоту ограничивается для пихты, ели, липы, ольхи черной 30 м, осины, березы – 35 м, дуба – 40 м, лиственницы, сосны – 45 м. Для сравнения пихта кавказская достигает 50 – 60 м, иногда – 80 м, ель кавказская 50 иногда 65 м.
Если принять предельную ширину окна в 1,5 высоты полога и длину в две ширины окна, оптимальные размеры окна составят (исключая кавказские виды) соответственно 0,4; 0,6; 0,7; 0,9 га. Приняв ориентировочно для главных лесообразователей средний верхний предел возраста насаждений ели – 250 лет, сосны – 350 лет и дуба 400 лет, что дает вместе с окнами число парцелл, соответственно 26, 36, 41, а среднее число видов вместе с сопутствующими 3; несложным расчетом получаем минимальный размер постоянного экологического участка, равный для ели 0,6 ∙ 26 ∙ 3 = 47 га, сосны 0,9 ∙ 36 ∙ 3 = 97 га, дуба 0,7 ∙ 41 ∙ 3 = 86 га.
Колебания давления среды на каждую особь происходят, как правило, циклически. Так, по данным Н. Я. Саликова, полноты древостоев колеблются от критических самоизреживающизся 0,67 до максимальных предельно плотных 1,33. Колебания давления среды связаны с увеличением объема пространства дерева во время роста. С учетом микроколебаний, богатства почвы, разной энергии роста деревьев, некомпактного (по сравнению с компактным равносторонним треугольным расположением) размещения по площади, выпадения отставших в росте, что освобождает ресурсы для оставшихся, давление среды всегда неодинаковое для отдельно рассматриваемых особей. До определенного возраста (примерно 100 лет) отпад положительно влияет на рост и развитие древостоя. Это этап динамических изменений – сукцессия, происходящая непрерывно, но с большими изменениями среды в первоначальном периоде от стадии окна и постепенным замедлением изменений, примерно, до 100 лет, после чего насаждение входит в этап гомеостаза. Ю. Одум (1980 г.) под экологической сукцессией понимает вообще весь процесс развития экосистемы. Более конкретное определение дает этому явлению Н. Ф. Реймерс (1990 г.): «Сукцессия – последовательная смена биогеоценозов, приемственно возникающая на одной и той же территории (биотопе) под влиянием природных факторов (в том числе и внутренних противоречий самих биоценозов) или воздействия человека». Изменения в сообществе в результате сукцессии носят закономерный характер и обусловлены взаимодействием организмов между собой и с окружающей абиотической средой.
Экологическая сукцессия происходит в определенный отрезок времени, в который изменяется видовая структура сообщества и абиотическая среда его существования вплоть до кульминации его развития – возникновения стабилизированной системы. Такую стабилизированную экосистему называют климаксом. В этом состоянии система находится тогда, когда в ней на единицу энергии приходится максимальная биомасса и максимальное количество симбиотических связей между организмами (Ю. Одум, 1975 г.). Однако к этому состоянию система проходит через ряд стадий развития, первые из которых часто называют стадией первых поселенцев. Поэтому, в более узком смысле, сукцессия – это последовательность сообществ, сменяющих друг друга в данном районе.
Стабильность сообщества может быть длительной лишь в том случае, если изменения среды, вызванные одними организмами, точно компенсируются деятельностью других, с противоположными экологическими требованиями. Это условие нарушается при нарушении круговорота веществ и тогда часть популяций, которые не могут выдержать конкуренции, вытесняются другими, для которых эти условия благоприятны, и гомеостаз восстанавливается.
Для возникновения сукцессии необходимо свободное пространство. В зависимости от первоначального состояния субстрата, различают первичную и вторичную сукцессии. Первичная сукцессия – это если формирование сообществ начинается на первоначально свободном субстрате, а вторичная сукцессия – это последовательная смена одного сообщества, существовавшего на данном субстрате, другим, более совершенным для данных абиотических условий.
Первичная сукцессия позволяет проследить формирование сообществ с самого начала. Она может возникнуть на склоне после оползня или обвала, на образовавшейся отмели при отступлении моря и изменении русла рекой, на обнаженных эоловых песках пустыни, не говоря уже об антропогенных нарушениях: свежая лесосека, намывная полоса морского побережья, искусственные водохранилища.
Первыми, как правило, на свободное пространство начинают внедряться растения посредством перенесенных ветром спор и семян, либо за счет вегетативных органов, оставшихся по соседству растений. В качестве примера первичной сукцессии можно привести зарастание еловым лесом новых территорий на севере России.
Ельник – это последняя климаксная стадия развития экосистемы в климатических условиях Севера, т. е. уже коренной биоценоз. Вначале же здесь развиваются березняки, ольховники, осинники, под пологом которых растут ели. Постепенно они перерастают березу и вытесняют ее, захватывая пространство. Семена обеих древесных пород легко переносятся ветром, но, если даже они прорастут одновременно, береза растет намного быстрее – к 6 – 10 годам ель едва достигает 50 – 60 см, а береза – 8 – 10 м. Под уже сомкнутыми кронами берез возникает свой микроклимат, обилие опада листьев способствует формированию особых почв, поселяются многие животные, появляется разнообразный травянистый покров, создаются консорции березы с окружающей средой. А ель продолжает расти в столь благоприятной обстановке и, наконец, береза не выдерживает конкуренции с ней за пространство и свет и вытесняется елью.
Классическим примером природной сукцессии является «старение» озерных экосистем – эвтрофикация. Она выражается в зарастании озер растениями от берегов к центру. Здесь наблюдается ряд стадий зарастания – от начальных – дальние от берега до достигнутых у берега. В конечном итоге озеро превращается в торфяное болото, представляющее собой устойчивую экосистему климаксного типа. Но и она не вечна – на ее месте постепенно может возникнуть лесная экосистема уже благодаря наземной сукцессионной серии в соответствии с климатическими условиями местности.
Эвтрофикация водоема в значительной степени определяется привносом извне биогенных элементов. В природных условиях биогены сносятся с площади водосбора. Такая эвтрофикация имеет черты первичной прогрессивной сукцессии.
Вторичная сукцессия является, как правило, следствием деятельности человека. В частности, описанная выше смена растительности при формировании ельника чаще происходит в результате вторичной сукцессии, возникающей на вырубках ранее существовавшего леса (ельника). Вторичная сукцессия заканчивается стабильной стадией сообщества через 150 – 250 лет, а первичная длится 1000 лет.
Вторичная, антропогенная сукцессия проявляется так же и в эвтрофикации. Бурное «цветение» водоемов, особенно искусственных водохранилищ, есть результат их обогащения биогенами, обусловленное деятельность человека. «Пусковым механизмом» процесса обычно является обильное поступление фосфора, реже – азота, иногда углерода и кремния. Ключевую роль обычно играет фосфор.
При поступлении биогенов резко возрастает продуктивность водоемов за счет роста численности и биомассы водорослей, и прежде всего сине-зеленых – цианей, из царства дробянок. Многие из них могут фиксировать молекулярный азот из атмосферы, тем самым снижая лимитирующее действие азота, а некоторые способны освобождать фосфор из продуктов метаболизма различных водорослей. Обладая этим и рядом других подобных качеств, они захватывают водоем и доминируют в биоценозе.
Биоценоз практически полностью перерождается. Наблюдаются массовые заморы рыб. «В особо тяжелых случаях вода приобретает цвет и консистенцию горохового супа, неприятный гнилостный запах: жизнь аэробных организмов исключена» (Соловьев, 1987 г.).
Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называется сукцессионной серией. Она наблюдается в природе не только в лесах, болотах и озерах, но и на стволах отмирающих деревьев и в пнях, где происходит закономерная смена сапрофитов и сапрофагов, в лужах и прудах и т.д. Иными словами сукцессии разномасштабны и иерархичны, так же как и сами экосистемы.
Гомеостаз – способность биологических систем – организма, популяции и экосистем – противостоять изменениям и сохранять равновесие. Исходя из кибернетической природы экосистем – гомеостатический механизм – это обратная связь. Например, у пойкилотермных животных изменение температуры тела регулируется специальным центром в мозге, куда постоянно поступает сигнал обратной связи, содержащий данные об отклонении от нормы, а от центра поступает сигнал, возвращающий температуру к норме. В механических системах аналогичный механизм называют сервомеханизмом, например, термостат управляет печью.
Для управления экосистемами не требуется регуляция извне – это саморегулирующаяся система. Саморегулирующий гомеостаз на экосистемном уровне обеспечен множеством управляющих механизмов. Один из них – субсистема «хищник – жертва» (рис. 5).

Рис. 5. Элементы кибернетики (по Ю. Одуму, 1975 г. с изменениями): a – взаимодействие положительной (+) и отрицательной (-) обратных связей в системе хищник – жертва.
Между условно выделенными кибернетическими блоками управление осуществляется посредством положительных и отрицательных связей. Положительная обратная связь «усиливает отклонение», например, увеличивает чрезмерно популяцию жертвы. Отрицательная обратная связь «уменьшает отклонение», например, ограничивает рост популяции жертвы за счет увеличения численности популяции хищников. Эта кибернетическая схема отлично иллюстрирует процесс коэволюции в системе «хищник-жертва», так как в этой «связке» развиваются и взаимные адаптационные процессы. Если в эту систему не вмешиваются другие факторы (например, человек уничтожил хищника), то результат саморегуляции будет описываться гомеостатическим плато (рис. 6) – областью отрицательных связей, а при нарушении системы начинают преобладать обратные положительные связи, что может привести к гибели системы.
Применительно к лесному фитоценозу гомеостаз характеризуется относительно постоянным числом эдификаторов и упрочением связей между всеми ярусами и видами, входящими в биогеоценоз, вселение «чужаков» в сообщество невозможно, т. к. не имеется свободных ресурсов. Выпадение от модификационных факторов отдельных деревьев верхнего яруса приводит к появлению просвета в пологе, так называемому «ветровально – почвенному комплексу», в виде гантели на месте упавшей кроны и выворота корней. Освободившиеся ресурсы, как правило, заполняются групповым подростом от произрастающих в биогеоценозе видов. Постепенно просветы расширяются до размеров окон. Климаксовый лес всегда неоднороден, в нем имеются многочисленные экологические ниши в виде захламленности, микровозвышений и понижений, разной освещенности, муравьиных куч, кротовин и т. д., что увеличивает биоразнообразие, а значит и устойчивость леса. В целом прирост биомассы климаксового биогеоценоза равен нулю, т. е. в одной части идет нарастание прироста, в другой – падение, в третьей наблюдается равновесие.
Наиболее устойчивы крупные экосистемы, и самая стабильная из них - биосфера, а наиболее неустойчивы – молодые экосистемы. Это объясняется тем, что в больших экосистемах создается саморегулирующий гомеостаз за счет взаимодействия круговоротов веществ и потоков энергии (Ю. Одум, 1975 г.).
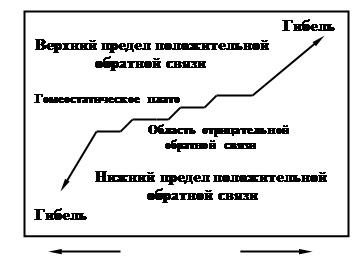
Рис. 6. Представление о гомеостатическом плато, в пределах которого поддерживается относительное постоянство вопреки условиям, вызывающим отклонения.
Вопрос № 28
Концепция климаксового леса: понятие, внутренняя и внешняя структуры, связь с экотопом. Влияние антропогенных факторов на лес.
В предыдущем вопросе при рассмотрении сукцессионных процессов мы уже дали определение климакса, как стабилизированной экосистемы. Первые переселенцы, которые приживаются на новом участке, - это организмы, которые толерантны к абиотическим условиям нового для них местообитания. Не встречая особого сопротивления среды, они чрезвычайно быстро размножаются (саранча, эфемерная растительность и т.п.), т. е. на ранних этапах в эволюции экосистемы преобладает г-стратегия (рост численности). Но постепенно возрастает видовое разнообразие за счет достаточно быстрой смены и увеличения количества популяций и начинает возрастать значение К-фактора (ограничитель роста).
Увеличение видового разнообразия приводит к усложнению связей внутри сообщества, умножению симбиотических связей, снижению чрезмерной рождаемости и доминирования массовых видов, и т. д. Наконец, действия г- и К-факторов уравновешиваются и сообщество развивающейся серии становится стабильным, или климаксным, - «это самоподдерживающееся сообщество, находящееся в равновесии с физическим местообитанием» (Ю. Одум, 1975 г.). Развивающееся сообщество преобразует и само местообитание.
На первых этапах для растительных форм первостепенное значение имеют почвенные биогенные элементы. Но черпать их из запасов почв до бесконечности невозможно и по мере истощения этих запасов разложение отмершей органики становится основным источником питания минеральными веществами биогеохимического круговорота.
Однако такой круговорот возможен лишь в автотрофной системе, черпающей энергию от солнца. Другое дело – гетеротрофная сукцессия, когда приток мертвого органического вещества не восполняет запасы, т. е. первичная продукция равна нулю, и участвуют в сукцессии только гетеротрофные организмы. В этом случае количество энергии не добавляется, а уменьшается, и система прекращает свое существование – все организмы погибают или, в лучшем случае, переходят в покоящиеся стадии. Характерным примером такой сукцессии является сукцессия в гниющих стволах деревьев, в трупах животных, фекалиях и на вторичных стадиях обработки сточных вод. Такая модель сукцессии должна ассоциироваться, по мнению Ю. Одума (1975 г.), с эксплуатацией залежей горючих полезных ископаемых человеком.
На ранних стадиях сукцессионной серии чистой продукции получается значительно больше и при ее изымании человеком сукцессия только приостанавливается, но основа продуктивности на этих этапах не подрывается. Другое дело в климаксных сериях – здесь чистая продуктивность снижается и, в принципе, становится константой. В этом случае очень важно знать величину этой константы с тем, чтобы четко представлять себе ту величину чистой продукции, которую можно изъять из системы, сохранив ее способность к самовосстановлению.
Так, например, вырубку лесов надо вести на локальных участках, с оставлением части территории с коренными типами пород. Это сократит время восстановления фитоценозов, так как сукцессионные серии сократятся до нескольких десятилетий (30 – 50 лет). Чтобы получить полноценную популяцию сосны, состоящую из 30 мозаик от 10-летней биогруппы до 300-летней, необходим период в 300 лет, а чтобы превратить разновозрастную популяцию в одновозрастную рубкой главного пользования достаточно одного года. Сплошная рубка приведет к разрушению всей экосистемы, в том числе ее эдафической части. Это приведет к нарушению сложившейся водонакопительной системы обширного региона, сменит лесной режим, переводящий осадки во внутрипочвенный сток, повлияет на режим открытого места, с резкими колебаниями температур и поверхностным стоком воды. Вырубка переходного типа растительности приводит к появлению пустоши, десятилетиями не зарастающей лесом, а рубка на крутых склонах приводит к потере лесной площади навсегда. Восстановление лишь почв потребует тысячелетия. Более того, сукцессионная серия может пойти и не по пути формирования прежнего лесного сообщества, а, скажем, по пути формирования пустыни и болот или других малопродуктивных экосистем. По теории островной биогеографии (А. В. Яблоков, С. А. Остроумов, 1983 г.), массив леса менее 250 тысяч га не способен обеспечить полноценную жизнь всем лесным видам, неизбежна гибель многих из них из-за недостатка территории для существования минимально необходимого числа особей в популяции.
Таким образом, сообщество не может одновременно быть высокостабильным и давать большой выход чистой продукции, который можно было бы изъять без вреда для самого биоценоза.
В почвенной биоте столь же активно протекают сукцессионные процессы. Они обусловлены разложением органического вещества и лежат в основе биологических круговоротов, - естественных регуляторов процессов, обеспечивающих плодородие почвы. Загрязнение почвенной среды и нарушение процессов образования гумуса снижают регуляторную способность почв и ведут к подрыву естественного плодородия, а следовательно, и к изменениям в экосистеме. Таким образом, эдафическая компонента может весьма существенно повлиять на ход экологической сукцессии при нарушении ее регуляторной функции.
Полнота сукцессий и видовое разнообразие возможны в случае надежной «работы» круговорота питательных веществ. Только в этом случае можно говорить о стабильности экосистемы, которая достигается в результате преобразования сообщества на основе длительной эволюции видов.
Во избежание экологической катастрофы сельхозугодия и урбанизированные территории (дороги, промышленность, постройки и т. д.) не должны занимать более 1/3 суши, 1/3 должна сохраняться естественно и 1/3 может быть частично изменена хозяйственной деятельностью человека. При этом деятельность должна быть разумной, нельзя, например, снижать полноту ниже 0,67 во избежание возникновения морозобойной ямы из-за неспособности верхнего полога удержать ночью накопившееся за день тепло. Перевод высоковозрастного климаксового леса, где не более 30% территории занято молодняками и средневозрастными, в неустойчивые динамические леса, где самые старые парцеллы не старше 120 лет, высвобождает огромное количество углекислого газа, создающего парниковый эффект в планетарном масштабе со всеми его негативными последствиями. Перечисленного ниже краткого перечня мероприятий достаточно, чтобы понять острейшую необходимость восстановления внешней структуры леса. Возможны следующие мероприятия при недостаточном финансировании:
· отвод площадей по водоразделам под лесозаращивание для соединения отдельных колков леса, с мерами содействия естественному возобновлению;
· переход на котловинные и группово-постепенные рубки в виде окон;
· замена рубок ухода по всей площади на рубки биогруппами;
· оставление резерватов (не менее 10% в квартале) на вырубаемых площадях, размерами 200 x 200 м для сохранения биоразнообразия лесных видов растений и животных;
· выделение особо защитных участков (ОЗУ) – «памятников природы» - на границах распространения ареалов видов, например, лиственницы, пихты, ясеня и т. д., что позволит сохранить генетический фонд и биоразнообразие;
· исключить из расчета главного пользования переходные типы растительности;
· искусственное создание окон путем вырубки для восстановления биоразнообразия мозаично-ярусной структуры в насаждениях всех возрастов. На бедных почвах возможна сплошная вырубка окнами, а на богатых – постепенная – выборочными рубками.
Полным биологическим разнообразием обладает биосфера, которая и является самой стабильной глобальной экосистемой – экосферой. Но биологическое разнообразие, обеспечивающее ее стабильность, - это прежде всего разнообразие стабильных природных экосистем, отличающихся видовым разнообразием естественной биоты.
Вопрос № 38
Математическое моделирование в экологии и в охране окружающей среды.
Системный подход в экологии обусловил формирование целого направления, ставшего ее самостоятельной отраслью – системной экологией. Системный подход – это направление в методологии познания объектов как систем. Система – это множество взаимосвязанных элементов, образующих определенную целостность, единство. Ее состав, структуру и свойства изучают посредством системного анализа, являющегося основой системного подхода и представляющего собой совокупность методологических средств, используемых для решения сложных научных проблем. В эту совокупность средств входит комплекс методов от простых описательных, логических до весьма сложных математических. Технической основой системного анализа являются современные ЭВМ и информационные системы с широким использованием методов математического программирования, теории игр и т.д.
Основными системными принципами являются: целостность, структурность, взаимозависимость системы и среды, иерархичность, множественность описания каждой системы. Целостность – обобщенная характеристика системы, свойства которой несводимы к сумме свойств ее элементов и не выводимы из этих свойств (целостность организмов более полной будет в популяции, популяции – в биоценозе и т. д., и свойства каждой системы не сводимы к свойствам нижестоящих). Структурность – установление структуры и взаимозависимости структурных элементов, обусловленности поведения системы ее структурой (структура биоценоза, трофическая структура экосистемы в установление измеримых связей между трофическими уровнями, и др.). Взаимозависимость системы и среды выражается в формировании и проявлении ее свойств в результате этого взаимодействия (взаимодействие биоценоза и биотопа, популяций в биоценозе и т.п.). Иерархичность – это когда каждый компонент системы может рассматриваться как самостоятельная система, а сама исследуемая система является составной частью более широкой системы (уровни биологической организации, вплоть до глобальной системы – биосферы).
Экосистемы – это весьма сложные самоорганизующиеся и целенаправленные, со сложной иерархической структурой системы, требующие множественного описания каждой системы, что требует построения множества моделей, т. е. широкого использования методов моделирования при исследовании.
Построение обобщенных моделей, отражающих все факторы и взаимосвязи в системе, является центральной процедурой системного анализа. Понятие «модель» широко используется, например, на бытовом уровне: модель самолетов, кораблей, автомобилей и т. п. Если эти модели не действующие, то они отражают только морфологические особенности объекта, но уже знание этих особенностей позволяет человеку, если он раньше не видел оригинал, узнать этот оригинал по модели. Иными словами, лишь часть свойств объекта позволяют судить об объекте в целом, в данном случае – о форме объекта. Нечто похожее происходит и при научных исследованиях.
Традиционная схема научного исследования: исследователь – объект. Здесь исследователь получает информацию путем непосредственного изучения объекта. Например, биолог изучает видовой состав фитопланктона под микроскопом. Но такое возможно лишь на достаточно простых объектах, но не при исследовании целостной структуры экосистемы, взаимодействия ее компонентов и т. п. В этом случае необходимо моделирование, при котором работает схема: исследователь – модель – объект изучения.
Например, чтобы получить представление об энергетических потоках в экосистеме, необходимо представить себе модель в виде пирамиды энергий или хотя бы пирамиды Элтона и т. п. Здесь появляется промежуточный (вспомогательный) объект изучения – модель.
Модель – это вспомогательный объект, находящийся в определенном объективном соответствии с познаваемым оригиналом и способный замещать его на отдельных этапах познания. Моделирование – это разработка, исследование модели и распространение модельной информации на оригинал (Лиепа, 1982 г.). Достоинства моделирования проявляются там, где возможности традиционного подхода оказываются ограниченными. Именно такой областью познания является экология.
Модель должна соответствовать двум требованиям: 1) она должна отражать лишь те особенности оригинала, которые выступают в качестве предмета познания, и 2) она должна быть адекватна оригиналу (иначе представления о нем будут искажены). Сам процесс моделирования, по И. Я. Лиепа (1982 г.), можно разделить на четыре этапа: качественный анализ, математическая реализация, верификация и изучение моделей.
Первый этап моделирования – качественный анализ - является основой любого объектного моделирования. На его основе формируются задачи, и выбирается вид модели. Этот этап обязан обеспечить соответствие модели двум вышеуказанным требованиям. Вид модели выбирается исходя из способа построения, из характера самого объекта и др.
По способу построения все модели делят на два класса: материальные и абстрактные. Материальные модели по своей физической природе сходны с оригиналом. Они могут сохранять геометрическое подобие оригиналу (макеты, тренажеры, искусственные заменители органов и т. д.), подобие протекания физических процессов с оригиналом – физическое моделирование (гидрологическая модель – течение воды и т. п.) и могут быть природными объектами – прообразами оригинала, т. е. натурными моделями (метод пробных участков). Материальные модели используются обычно в технических целях и мало подходят для экологических проблем. Более подходящими для экологического моделирования являются абстрактные модели, представляющие собой описание оригинала в словесной форме или посредством символов и операций над ними, отражающих исследуемые особенности оригинала. Абстрактные модели подразделяются на три типа: вербальные, схематические и математические.
Вербальные модели – это формализованный вариант традиционного естественнонаучного описания в виде текста, таблиц и иллюстраций (Федоров, Гильманов, 1980 г.). Схематические модели разрабатываются в виде различного рода схем, рисунков, графиков и фотографий, основные их достоинства – наглядность, информативность и простота построения (трофические цепи, пирамида Элтона, схемы структуры, динамики и энергетики экосистем, воздействия экологических факторов, биохимических круговоротов, и др.).
Вербальные и схематические модели – неотъемлемая часть качественного анализа математического моделирования, являющегося наиболее совершенным видом количественного исследования оригинала, позволяющая построить его математическую модель. «Математическая модель» - это математическое описание оригинала, отражающее его целостность, структуру, динамику, функционирование и взаимосвязи оригинала, внешних и внутренних факторов воздействия» (Лиепа, 1982 г.). Это означает, что практически такая модель есть формула или система уравнений и неравенств.
По своему характеру выделяют модели статические и динамические. Статическая модель отражает объект (систему), не изменяющий свое состояние во времени, а динамическая модель отражает объект (систему), изменяющий свое состояние во времени. Подавляющее большинство живых объектов и систем – это динамические системы и могут быть отражены только лишь динамическими моделями.
Второй этап моделирования – это математическая реализация логической структуры модели. С точки зрения технологии применения математических методов можно выделить модели аналитические и численные (компьютерные). Аналитическая модель – это построение теоретических концепций с применением строгого математического аппарата, обычно позволяющего вывести общую формальную зависимость. Компьютерные модели П. М. Брусиловский, Г. С. Розенберг (1981 г.) делят на имитационные и самоорганизующиеся.
Имитационные модели отражают представления исследователя о взаимосвязях в экосистеме и как они реализуются. Наилучшие результаты эти модели дают при составлении прогноза изменений в экосистеме. Самоорганизующиеся модели относятся к классу регрессионных уравнений, в них широко используются вероятностно статистические методы расчетов.
Третий этап моделирования предусматривает верификацию модели: проверку соответствия модели оригиналу. На данном этапе необходимо удостовериться, что выбранная модель отвечает второму требованию: адекватно отражает особенности оригинала. Для этого может быть проведена эмпирическая проверка – сравнение полученных данных с результатами наблюдений за оригиналом. Модель может быть признана высококачественной, если прогнозы оправдываются. При отсутствии эмпирических данных проводится теоретическая верификация – по теоретическим представлениям определяется область применения и прогностические возможности модели.
Четвертый этап моделирования – это изучение модели, экспериментирование с моделью и экологическая интерпретация модельной информации. Основная часть этапа – выявление новых закономерностей и исследование возможностей оптимизации структуры и управление поведением моделируемой системы, а также пригодность модели для прогнозирования.
В экологии математические модели экосистем В. Д. Федоров и Т. Г. Гильманов (1980 г.) предлагают разделить на модели популяционного, биоценотического и экосистемного уровней. Популяционные модели описывают особенности отдельных популяций, отражают их свойства и внутренние закономерности: модели, позволяющие оценить динамику численности и возрастного состава популяций в зависимости от рождаемости и смертности, заданных как функции лишь от общей плотности и возрастного состава популяций. Модели биоценотического уровня задаются, как системы уравнений, отражающих динамику биоценоза как функцию плотностей составляющих его популяций. Модели экосистемного уровня представляют собой системы уравнений, в число аргументов которых включены как внутренние переменные состояния, так и внешние факторы воздействия и целостные свойства экосистем. Модели данного уровня учитывают и роль обратных связей в функционировании систем.
При построении любой модели главная задача – создать модель достаточной полноты. Для этого необходимо стремиться учесть все существенные факторы, влияющие на рассматриваемые явления; уделить специальное внимание наличию в ней противоречивых элементов, как одного из признаков полноты модели; учесть возможность появления неизвестных факторов, чтобы в случае необходимости дополнить модель новым элементом.
Биология – одна из первых наук, в которой приоритетное значение приобрел системный подход в изучении природы, впервые в научной форме использованный Ч. Дарвином. Особенно широко используются системные идеи в экологии. На новую, более высокую ступень идеи системного подхода поставлены в учении В. И. Вернадского о биосфере и ноосфере, где научному познанию предложен новый тип объектов – глобальные системы. Такой глобальной экосистемой и является биосфера, объединяющая на основе иерархического принципа все экосистемы Земли более низких уровней.
Вопрос № 48
Организационная структура системы управления, регулирования, охраны и контроля окружающей среды и природопользования в стране и области. Экологическая экспертиза. Критерии качества окружающей среды. Экологические подходы к нормированию предельно допустимых концентраций, выбросов, нагрузок, сбросов (ПДК, ПДВ, ПДН, ПДС).
Государственные органы управления, контроля и надзора в области охраны окружающей природной среды подразделяются на две категории: органы общей и специальной компетенции.
К государственным органам общей компетенции относится Президент, Федеральное Собрание, Государственная Дума, Правительство, представительные и исполнительные органы власти субъектов Российской Федерации, муниципальные органы. Эти органы определяют основные направления природоохранной политики, утверждают экологические программы, обеспечивают экологическую безопасность, устанавливают правовые основы и нормы в пределах своей компетенции, и т. д. Наряду с охраной окружающей природной среды государственные органы этой категории ведают и другими экологическими вопросами, входящими в круг их полномочий.
Государственные органы категории специальной компетенции подразделяются на комплексные, отраслевые и функциональные.
Комплексные органы выполняют все природоохранные задачи или какой-либо их блок. До 2000 г. функция управления природопользованием и охраны окружающей природной среды выполняли Государственный комитет РФ по охране окружающей среды (Госкомэкологии России) и Министерство природных ресурсов РФ (МПР России) и другие ведомства. Указом Президента РФ от 17 мая 2000 г. Государственный комитет РФ по охране окружающей природной среды был упразднен и его функции переданы Министерству природных ресурсов РФ, в структуру которого вошли Государственная служба охраны окружающей среды и Государственная служба контроля в сфере природопользования и экологической безопасности.
К комплексным органам относятся также:
· Департамент Госсанэпиднадзора Министерства РФ (Санэпиднадзор РФ) – координатор деятельности всех ведомств и организаций в области санитарной охраны через систему территориальных органов (санэпидстанций и инспекций);
· ошли Государственная служба охраны окружающей среды и Государственная служба контроля в сфере природопользования и экологической безопасности.
К комплексным органам относятся также:
· Департамент Госсанэпиднадзора Министерства РФ (Санэпиднадзор РФ) – координатор деятельности всех ведомств и организаций в области санитарной охраны через систему территориальных органов (санэпидстанций и инспекций);
· Федеральная служба России по гидрометеорологии и мониторингу окружающей среды (Росгидромет) – осуществляет экологический контроль за состоянием окружающей природной среды, информирует население об изменениях в окружающей среде с помощью широкой сети наблюдательных пунктов и др.;
· Министерство РФ по делам гражданской обороны, чрезвычайных ситуаций и ликвидации последствий стихийных бедствий (МЧС России) – обеспечивает безопасность людей в условиях экстремальной ситуации, стихийных бедствий, производственных аварий и катастроф.
Отраслевые органы (Роскомзем, Рослесхоз, Госкомрыболовство, Минсельхоз России) выполняют функции управления и надзора по охране и использованию отдельных видов природных ресурсов и объектов.
Функциональные органы выполняют одну или несколько родственных функций в отношении природных объектов: Минатом России (обеспечение ядерной и радиационной безопасности); Госгортехнадзор России (контроль за использованием недр); Минздрав России (санитарно-эпидемиологический контроль); МВД России (охрана атмосферного воздуха от загрязнения транспортными средствами, санитарно-экологическая служба муниципальной милиции).
Согласно статье 9 Закона РФ «Об охране окружающей природной среды» в компетенцию краев, областей в сфере охраны окружающей природной среды входят следующие мероприятия:
· определение основных направлений охраны окружающей природной среды и утверждение территориальных экологических программ;
· учет и оценка природной среды, состояния природных ресурсов, учет экологически вредных объектов, ведение кадастровой документации;
· учет и оценка объемов образующихся отходов производства и отработанных продуктов на предприятиях, в учреждениях и организациях, расположенных на территории региона, независимо от их форм собственности и подчиненности;
· планирование охраны окружающей природной среды, финансирование и материально-техническое обеспечение природоохранных программ;
· координация природоохранительной деятельности органов государственного управления, предприятий, учреждений, организаций, содействие добровольному кооперированию средств на выполнение мероприятий по охране окружающей природной среды;
· координация деятельности экологических служб предприятий, учреждений, организаций независимо от форм собственности и подчинения, проведение экологической экспертизы проектов;
· государственный экологический контроль, принятие решений об ограничении, приостановлении, прекращении деятельности объектов, не отвечающих требованиям природоохранительного законодательства;
· запрещение строительства экологически вредных объектов;
· дача разрешений на право использования природной среды и ее ресурсов, на выброс и сброс вредных веществ, размещение, переработку, складирование и захоронение отходов;
· организация сбора и рекуперация производственных и бытовых отходов;
· организация особо охраняемых природных территорий;
· организация экологического воспитания, образования и просвещения;
· обеспечение населения необходимой экологической информацией.
Правовой механизм управления природопользованием и охраной окружающей природной среды включает в себя и такую важную форму предупредительного экологического контроля, как экспертизу. Различают государственную, общественную и иные виды экологических экспертиз.
Под государственной экологической экспертизой понимается проверка представленных материалов специальной комиссией, называемой Госкомэкологии России. Задача экспертной комиссии – оценить соответствие намечаемой хозяйственной или иной деятельности требованиям экологической безопасности.
Объектами государственной экологической экспертизы являются любые проектные и предпроектные документы, новая техника и технология, продукция, сырье и материалы, вещества, а также проекты стандартов и нормативов.
Федеральным законом «Об экологической экспертизе» (1995 г.) установлены следующие принципы государственной экологической экспертизы: обязательность ее проведения, научная обоснованность выводов, независимость и вневедомственность – широкая гласность, привлечение общественности, а главное, презумпция потенциальной экологической опасности любой намечаемой хозяйственной и иной деятельности.
Государственная экологическая экспертиза, как правило, предшествует принятию хозяйственного решения. Это позволяет еще на стадии планирования и проектирования выявить допущенные ошибки, оценить их последствия и дать рекомендации по их устранению. Финансирование работ по всем проектам и программам открывается только при наличии положительного заключения государственной экологической экспертизы.
Кроме государственной существуют и другие виды экспертиз – общественная, научная и другие, которые проводятся обычно на добровольной основе и носят рекомендательный или информационный характер.
Правовой основой экологической экспертизы служит Федеральный закон «Об экологической экспертизе» (1995 г.), а также постановления, указы и другие природоохранные акты. Нормативной базой являются стандарты, нормы, правила и т. д., обобщенные в специальных справочниках для экспертов.
Об эффективности государственной экологической экспертизы свидетельствует тот факт, что только в 1997 году из 55 тысяч рассмотренных материалов было отклонено и отправлено на доработку около 15 тысяч.
Экологическая экспертиза становится одной из важных функций государственной экологической политики. Сейчас уже невозможно представить превентивное правовое регулирование хозяйственной деятельности без экологической экспертизы, нацеленной на снижение экологического риска при принятии решений. Еще более глубоким и объемным вариантом проведения экологической экспертизы в последние годы служит – оценка воздействия на окружающую среду (ОВОС). Здесь учитывается уже и трансграничный перенос загрязняющих веществ, информации, энергии и др.
Под качеством окружающей природной среды понимается степень соответствия ее характеристик потребностям людей и технологическим требованиям. В основу всех природоохранных мероприятий положен принцип нормирования качества окружающей природной среды. Этот термин означает установление нормативов (показателей) предельно допустимых воздействий человека на окружающую природную среду.
Согласно природоохранному закону Российской Федерации (1991 г.) соблюдение экологических нормативов, т. е. нормативов, которые определяют качество природной среды, обеспечивает:
· экологическую безопасность населения;
· сохранение генетического фонда человека, растений и животных;
· рациональное использование и воспроизводство природных ресурсов в условиях устойчивого развития.
Чем меньше пороговая величина экологических нормативов, тем выше качество окружающей природной среды. Однако более высокое качество требует соответственно больших затрат, эффективных технологий и высокочувствительных средств контроля. Поэтому нормативы качества окружающей природной среды по мере подъема уровня развития общества имеют тенденцию к ужесточению.
Основные экологические нормативы качества и воздействия на окружающую природную среду:
санитарно-гигиенические:
· предельно допустимая концентрация вредных веществ (ПДК);
· допустимый уровень физических воздействий (ПДУ) (шума, вибрации, ионизирующих излучений и др.);
производственно-хозяйственные:
· предельно допустимый выброс вредных веществ (ПДВ);
· предельно допустимый сброс вредных веществ (ПДС);
· допустимое изъятие компонентов природной среды;
· норматив образования отходов производства и потребления;
комплексные показатели:
· предельно допустимая антропогенная нагрузка на окружающую природную среду (ПДН).
Предельно допустимая концентрация (ПДК) – представляет собой количество загрязнителя в почве, воздушной или водной среде, которое при постоянном или временном воздействии на человека не влияет на его здоровье и не вызывает неблагоприятных последствий у его потомства. В последнее время при определении ПДК учитывается не только степень влияния загрязнения на здоровье человека, но и воздействие этих загрязнений на диких животных, растения, грибы, микроорганизмы, а также на природные сообщества в целом.
В настоящее время в нашей стране действуют более 1900 ПДК вредных химических веществ для водоемов, более 500 для атмосферного воздуха и более 130 для почв. ПДК устанавливают на основании комплексных исследований и постоянно контролируют органами гидрометеорологической службы Госкомсанэпиднадзора. ПДК не остаются постоянными, их периодически пересматривают и уточняют. После утверждения норматив становится юридически обязательным.
Для нормирования содержания вредного вещества в атмосферном воздухе установлены два норматива – разовый и среднесуточный ПДК. Максимально разовая предельно допустимая концентрация (ПДК м. р.) – это такая концентрация вредного вещества в воздухе, которая не должна вызывать при вдыхании его в течение 30 минут рефлекторных реакций в организме человека (ощущение запаха, изменение световой чувствительности глаз и др.). Среднесуточная предельно допустимая концентрация (ПДК с. с.) – это такая концентрация вредного вещества в воздухе, при которой на человека в течение неопределенно долгого (многолетнего) периода не оказывается прямое или косвенное вредное воздействие.
Значения ПДК наиболее часто встречающихся загрязнений атмосферного воздуха указаны в таблице 3.
При содержании в воздухе нескольких загрязняющих веществ, обладающих суммацией действия (синергизмом), например, диоксидов серы и азота; озона, диоксида азота и формальдегида, сумма их концентраций не должна превышать при расчете единицы:
С1/ПДК1 + С2/ПДК2 + … + Сn/ПДКn < 1,
где С1, С2, …, Сn – фактические концентрации вредных веществ в воздухе или воде; ПДК1, ПДК2, …, ПДКn – максимально разовые предельно допустимые концентрации вредных веществ, которые установлены для их изолированного присутствия, мг/м3.
Таблица 3
Предельно допустимые концентрации вредных веществ в атмосферном воздухе населенных пунктов, мг / м3
| Вещество |
Максимально разовая |
Среднесуточная |
| Азота оксид | 0,6 | 0,06 |
| Азота диоксид | 0,085 | 0,085 |
| Аммиак | 0,2 | 0,04 |
| Ацетон | 0,35 | 0,35 |
| Бензол | 1,5 | 0,8 |
| Бенз(а)пирен | - | 0,000001 |
| Пыль нетоксичная | 0,5 | 0,15 |
| Ртуть металлическая | - | 0,0003 |
| Сероводород | 0,008 | 0,008 |
| Сероуглерод | 0,03 | 0,005 |
| Углерода оксид | 5,0 | 5,0 |
| Фенол | 0,01 | 0,003 |
| Формальдегид | 0,035 | 0,003 |
| Хлор | 0,1 | 0,03 |
Под предельно допустимой концентрацией вредного вещества в почве (ПДК, мг/кг) понимается такая максимальная концентрация, которая не может вызвать прямого или косвенного влияния на среду, нарушить самоочищающую способность почвы и оказать отрицательное воздействие на здоровье человека.
Для водной среды ПДК загрязняющих веществ означает такую концентрацию этих веществ в воде, выше которой она становится непригодной для одного или нескольких видов водопользования. ПДК загрязняющих веществ устанавливаются отдельно для питьевых вод (таблица 4) и рыбохозяйственных водоемов.
Требования к качеству вод в водоемах, используемых для рыбохозяйственных целей, специфичны и в большинстве случаев более жестки, чем таковые для водных объектов хозяйственно-бытового назначения. Так, рыбохозяйственные ПДК для ряда моющих веществ в три раза ниже санитарных норм, нефтепродуктов – в шесть раз, а тяжелых металлов (цинка) – даже в сто раз. Объяснить это ужесточение требований к качеству воды в рыбохозяйственных водоемах не трудно, если вспомнить, что при переходе вредных веществ по пищевой (трофической) цепи происходит их биологическое накопление до опасных для жизни количеств.
Допустимый уровень радиационного воздействия на окружающую среду – это уровень, который не представляет опасности для здоровья человека, состояния животных, растений, их генетического фонда. ПДУ определяется на основании норм радиационной безопасности (НРБ – 76/87), основных санитарных правил (ОСП – 72/87) и санитарных норм проектирования(СП -254-71).
Установлены также предельно допустимые уровни (ПДУ) воздействия шума, вибрации, магнитных полей и иных вредных физических воздействий.
Допустимый выброс или сброс (ПДВ) и (ПДС) – это максимальное количество загрязняющих веществ, которые в единицу времени разрешается выбрасывать данному конкретному предприятию в атмосферу или сбрасывать в водоем, не вызывая при этом превышения в них предельно допустимых концентраций загрязняющих веществ и неблагоприятных экологических последствий.
Таблица 4
Предельно допустимые концентрации вредных веществ в питьевых водах, мг/л
| Вещество | ПДК | Вещество | ПДК |
| По санитарно-токсикологическому показателю | |||
| Анилин | 0,1 | Метаном | 3,0 |
| Бензол | 0,5 | Нитраты (по азоту) | 10,0 |
| Свинец (Pb+) | 0,1 | Ртуть | 0,005 |
| Стронций | 2,0 | Динитротолуол | 0,5 |
| Тетраэтилсвинец |
Отсутст- вует |
Формальдегид | 0,05 |
| Хлорбензол | 0,02 | Полиакриламид | 2,0 |
| По общесанитарному показателю | |||
| Аммиак (по азоту) | 2,0 | Метилпирролидон | 0,5 |
| Бутилацетат | 0,1 | Стрептоцид | 0,5 |
| Дибутилфталат | 0,2 | Тринитротолуол | 0,5 |
| Капролактам | 1,0 | ||
| По органолептическому показателю | |||
| Бензин | 0,1 | Диметилфенол | 0,25 |
| Бутилбензол | 0,1 | Динитробензол | 0,5 |
| Бутиловый спирт | 1,0 | Динитрохлорбензол | 0,5 |
| Гексахлоран | 0,02 | Дихлорметан | 7,5 |
Нормативами установлено, что если в воздухе городов или других населенных пунктов, где расположены предприятия, концентрации вредных веществ превышают ПДК, а значения допустимых выбросов по объективным причинам не могут быть достигнуты, вводится поэтапное снижение выброса вредных веществ до значений, обеспечивающих ПДК. При этом могут быть установлены временно согласованные выбросы (ВСВ) на уровне выбросов предприятий с наиболее совершенной или аналогичной ей технологией.
В настоящее время в России на нормативах допустимых выбросов работают лишь 15-20 % загрязняющих производств, на ВСВ (временно согласованных выбросах вредных веществ) – 40 – 50 %, а остальные загрязняют среду на основе лимитных выбросов и сбросов, которые определяют по фактическому выбросу на определенном отрезке времени.
Основным комплексным нормативом качества окружающей природной среды является допустимая норма антропогенной нагрузки.
Допустимые нормы антропогенной нагрузки на окружающую среду (ПДН) – это максимально возможные антропогенные воздействия на природные ресурсы или комплексы, не приводящие к нарушению устойчивости экологических систем.
Для оценки общей устойчивости экосистем к антропогенным воздействиям используют следующие показатели: 1) запасы живого и мертвого органического вещества; 2) эффективность образования органического вещества или продукции растительного покрова и 3) видовое и структурное разнообразие.
Ученые-экологи установили, что стабильность среды обитания не только растительного, но и животного мира, а в конечном счете и человека определяется в первую очередь, массой живого органического вещества и его основной части – фитомассы (древесина, травянистая растительность и др.). Чем значительнее эта масса, тем стабильнее среда. Главенствующее значение при этом имеют фотосинтезирующие организмы, так как они являются основным источником биомассы, а также определяют пищевые условия для всех остальных звеньев экосистемы и в значительной мере состав атмосферного воздуха.
Способность экосистем в минимальные сроки восстановиться в случае антропогенного нарушения определяется другим показателем – эффективностью образования продукции растительного покрова в результате вторичной сукцессии. Чем выше структурное и видовое разнообразие экосистемы, тем большее число комбинаций структурных элементов может создать она в ответ на внешнее антропогенное воздействие. Структурное разнообразие экосистемы можно оценить, сравнивая запасы фитомассы и зоомассы (хищники, копытные, грызуны и т. д.).
Потенциальная способность природной среды перенести ту или иную антропогенную нагрузку без нарушения основных функций экосистем определяется термином «емкость природной среды», или экологическая емкость территории.
Понятие о предельно допустимой антропогенной нагрузке на природную среду должно лежать в основе всего природопользования. В связи с этим различают экстенсивное и равновесное природопользование. Экстенсивное (расширяющееся) природопользование – это когда рост производства осуществляется за счет возрастающей нагрузки на природные комплексы, причем эта нагрузка растет быстрее, чем увеличивается масштаб производства. Экстенсивное природопользование может привести к полному разрушению природного комплекса и подобно тому, как безграничный рост биологической емкости популяции приводит к ее краху, крах потерпит и техносфера. Равновесное природопользование – это когда общество контролирует все стороны своего развития, добиваясь того, чтобы совокупная антропогенная нагрузка на среду не превышала самовосстановительного потенциала природных систем.
Отсюда вытекает важный вывод о том, что регулирование качества природной среды должно начинаться с определения нагрузок, допустимых с экологической точки зрения, а региональное природопользование должно соответствовать экологической «выносливости» территории.
Пренебрежение основным экологическим комплексным нормативом в инженерно-хозяйственной практике чревато серьезными экологическими просчетами. В 1990 году тогдашний руководитель Госкомприроды Н. Н. Воронцов отмечал: «Конечно же, нужно улучшать фильтры пыле- и газоуловителей, очищать сточные воды. Но не было у нас до сих пор главного – идеологии ресурсосбережения, определения экологической емкости, биосферного подхода».
При формировании территориально-производственных комплексов, развития промышленности, строительства, реконструкции городов и т. д. применение допустимых норм антропогенной нагрузки предусматривается в обязательном порядке. Региональные допустимые нормы антропогенной нагрузки устанавливают предельную хозяйственную нагрузку на территориальные природные комплексы. Отраслевые – на отдельные виды природных ресурсов, например, предельное число домашнего скота на единицу пастбищных угодий, предельное число посетителей в национальном парке и т. д.
Список литературы
1. Гальперин М. В. Экологические основы природопользования: Учебник. 2-е изд., испр. – М.: ФОРУМ: ИНФРА – М, 2005. – 256 с.: ил. – (Профессиональное образование).
2. Одум Ю. Экология / Пер. с англ. Т. 1 – 2. – М.: Мир, 1986. Стр. 328, 376.
3. Шамилева И. А. Экология: Учеб. пособие для студ. пед. вузов. – М.: Гуманит. изд. центр ВЛАДОС, 2004. – 144 с.: ил.
4. Арустамов Э. А., Баркалова Н. В., Левакова И. В. Экологические основы природопользования: Учебник. – 2-е изд. перераб. и доп./ Рук. авт. колл. Э. А. Арустамов. – М.: Издательско-торговая корпорация «Дашков и К`», 2005. – 320 с.
5. Коробкин В. И., Передельский Л. В. Экология / Изд. 8-е. – Ростов н/ Д. Феникс, 2005. – 576 с. (Высшее образование).
6. Степановских А. С. Охрана окружающей среды. Учебник для вузов / - М.: ЮНИТИ-ДАНА, 2000. – 559 с.
7. Татаринов К. П. Экология и охрана природы. Методические указания для студентов-заочников лесохозяйственного факультета специальности 260400/ НГСХА, Нижний Новгород, 2003. – 23 с.
Лимитирующие факторы
Впервые на значение лимитирующих факторов указал немецкий агрохимик Ю. Либих в середине XIX в. Он установил закон минимума: урожай (продукция) зависит от фактора, находящегося в минимуме. Если в почве полезные компоненты в целом представляют собой уравновешенную систему и только какое-то вещество, например, фосфор содержится в количествах, близких к минимуму, то это может снизить урожай. Но оказалось, что даже те же самые минеральные вещества, очень полезные при оптимальном содержании их в почве, снижают урожай, если они в избытке. Значит, факторы могут быть лимитирующими, находясь и в максимуме.
Таким образом, лимитирующими экологическими факторами следует называть такие факторы, которые ограничивают развитие организмов из-за недостатка или избытка по сравнению с потребностью (оптимальным содержанием). Их иногда называют ограничивающими факторами.
Что касается закона минимума Ю. Либиха, то он имеет ограниченное действие и только на уровне химических веществ. Р. Митчерлих показал, что урожай зависит от совокупного действия всех факторов жизни растений, включая температуру, влажность, освещенность и т. д.
Различия в совокупном и изолированном действиях относятся и к другим факторам. Например, действие отрицательных температур усиливается ветром и высокой влажностью воздуха, но, с другой стороны, высокая влажность ослабляет действие высоких температур, и т. д. Но, несмотря на взаимовлияние факторов, все-таки они не могут заменить друг друга, что и нашло отражение в законе независимости факторов В. Р. Вильямса: условия жизни равнозначны, но один из факторов жизни не может быть заменен другим. Например, нельзя действие влажности (воды) заменить действием углекислого газа или солнечного света, и т. д.
Наиболее полно и в наиболее общем виде всю сложность влияния экологических факторов на организм отражает закон толерантности В. Шелфорда: отсутствие или невозможность процветания определяется недостатком (в качественном или количественном смысле) или, наоборот, избытком любого из ряда факторов, уровень которых может оказаться близким к пределам переносимого данным организмом. Эти два предела называют пределами толерантности.
Относительно действия одного фактора можно проиллюстрировать этот закон так: некий организм способен существовать при температуре от – 5 о С до 25 о С, т. е. диапазон его толерантности лежит в пределах этих температур. Организмы, для жизни которых требуются условия, ограниченные узким диапазоном толерантности по величине температуры, называются стенотермными («стено» - узкий), а способных жить в широком диапазоне температур – эвритермными («эври» - широкий).
Подобно температуре действуют и другие лимитирующие факторы, а организмы по отношению к характеру их воздействия называют, соответственно, стенобионтами и эврибионтами. Например, говорят: организм стенобионтен по отношению к влажности, или эврибионтен к климатическим факторам, и т. п.
Диапазон толерантности организма не остается постоянным – он, например, сужается, если какой-либо из факторов близок к какому-либо пределу, или при размножении организма, когда многие факторы становятся лимитирующими. Значит, и характер действия экологических факторов при определенных условиях может меняться, т. е. он может быть, а может и не быть лимитирующим. При этом нельзя забывать, что организмы и сами способны снизить лимитирующее действие факторов, создав, например, определенный микроклимат (микросреду). Здесь возникает своеобразная компенсация факторов, которая наиболее эффективна на уровне сообществ, реже – на видовом уровне.
Такая компенсация факторов обычно создает условия для физиологической акклиматизации вида – эврибионта, имеющего широкое распространение, который, акклиматизируясь в данном конкретном месте, создает своеобразную популяцию, экотип, пределы толерантности которой соответствуют местным условиям. При более глубоких адаптационных процессах здесь могут появиться и генетические расы.
Итак, в природных условиях организмы зависят от состояния критических физических факторов, от содержания необходимых веществ и от диапазона толерантности самих организмов к этим и другим компонентам среды.