Курсовая работа: Общие черты предпатологии радиационной и нерадиационной природы
Курсовая работа
на тему:
"Общие черты предпатологии радиационной и нерадиационной природы"
2009
Введение
Наличие длительного скрытого периода развития радиационного лейкоза, так же как и других отдаленных радиационных последствий, заставило обратить паше внимание па более тщательное изучение того внешне клинически благополучного состояния организма, которое наступает вслед за практически полным восстановлением организма, перенесшего воздействие радиации. Радиационная модель, как известно, удобна для исследования общих закономерностей развития ряда предпатологических состояний вообще, так как клинические проявления и, по-видимому, патогенез отдаленной радиационной патологии принципиально не отличаются от патологии хронических радиационных и многих токсических воздействий. В частности, выяснилось, что в механизмах развития многих проявлений этих видов патологии важную роль играют ослабление и истощение одних и тех же компенсаторно-восстановительных реакций и искажение взаимосвязей различных систем организма, определяющих возникновение и функционирование так называемых порочных патогенетических кругов и имеющих связь с возрастными изменениями организма.
1. Радиационные изменения кроветворения в ближайшие и отдаленные сроки
При анализе процессов восстановления гематологических показателей у человека после острой лучевой болезни (средней степени тяжести) мы обратили внимание па то, что они восстанавливаются значительно позднее клинического выздоровления организма. Восстановление цитологического состава крови обычно наблюдается через 4-6 мес. и касается не всех ростков кроветворения.
В первую очередь восстанавливается красная кровь, если ее показатели были снижены. Восстановление числа лейкоцитов и тромбоцитов происходит в более поздние сроки и не носит характера устойчивой нормализации. Особенно долго не восстанавливается число лимфоцитов в крови. Так, через 1,5—2 года после радиационного воздействия еще находили пониженное содержание лимфоцитов в крови человека. Следует сказать, что восстановление цитологического состава крови часто сопровождается сохранением извращенных реакций белой крови па различные воздействия: инфекции, травмы, функциональные нагрузки.
Более подробно эти вопросы изучены в экспериментах. Так, в опытах И. Н. Усачевой последовательность восстановления различных ростков кроветворения у облученных собак была следующей. После воздействия рентгеновскими лучами в дозах 6 и 8 Гр через 45 сут. у выживших собак еще были значительные нарушения в периферической крови и костном мозге, несмотря на внешнее клиническое благополучие. Они выражались в уменьшении числа эритроцитов, тромбоцитов, лейкоцитов и лимфоцитов и в отсутствии нормального количественного соотношения эритробластических и миелоидных клеток костного мозга. Одновременно отмечалось значительное повышение числа ретикулоцитов с последующим нарастанием числа эритроцитов периферической крови и увеличением числа кроветворных клеток в костном мозге, среди которых преобладают эритробластические. Через 4—6 мес. у большинства собак наблюдалось первоочередное восстановление эритропоэза. Стойкого восстановления тромбопоэза не обнаруживали и через 18 мес. К этому сроку еще отмечали значительные колебания их числа в сторону как повышения, так и понижения. Еще медленнее шло восстановление белой крови. Даже через 12 мес. после облучения в дозе 7 Гр у четырех собак из десяти обследованных в этот срок число лейкоцитов было в пределах 3—6 тыс. клеток в 1 мм3 вместо исходных 9— 15 тыс. клеток. У двух собак, облученных в дозе 8 Гр и наблюдавшихся, и через 18—20 мес. число лейкоцитов не достигло исходного уровня. Эти изменения в значительной мере были обусловлены недостаточностью лимфопоэза.
Особый интерес представляют эксперименты Е.Н. Антипенко, специально поставленные для изучения последовательности восстановления показателей гемопоэза у собак после воздействия радиацией в дозах 4 и 6,5 Гр (часть собак при облучении в этих дозах подвергалась симптоматическому лечению). В группе выживших собак,
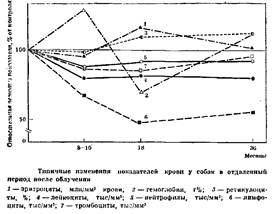
которых облучали в дозе 4,5 Гр (ЛД 50/30), восстановление гемопоэза происходило медленно. Быстрее всего восстанавливалось число эритроцитов. С 75-го дня после облучения их количество начинало превышать исходный уровень или равнялось ему. Число лейкоцитов за весь годовой период наблюдений не достигало исходных величин. Восстановление числа лимфоцитов шло наиболее замедленными темпами, и в последние месяцы эксперимента лимфоцитов в среднем оказалось в 2 раза ниже исходного уровня. Содержание тромбоцитов было значительно снижено — в большинстве случаев больше, чем нейтрофилов. Число тромбоцитов достигло уровня 90% исходной величины лишь на 12-м месяце. При исследовании костного мозга на 4-м и 10-м месяце отношение клеток миелоидного ряда к клеткам эритроидного ряда было смещено в сторону красного ряда.
Наблюдали значительное преобладание молодых форм клеток над зрелыми формами в нейтрофильном ряду, что указывало на сохранение в этом ряду сильной стимуляции к ускорению пролиферации клеток, способных к делению, в ущерб их созреванию.
Собаки, облученные в дозе 6 Гр с последующим симптоматическим лечением (в течение месяца), через 8— 10 мес. по общеклиническим показателям не отличались от здоровых животных. Исключение составили показатели крови и костного мозга (рис. 2). Во все сроки дальнейшего исследования (до трех лет после облучения) число лейкоцитов у них было снижено на 20%, так и не достигнув исходных величин. Это снижение происходило за счет еще большего уменьшения числа лимфоцитов — на 33—50%. В то же время показатели красной крови находились в пределах нормы или даже несколько превышали ее. Следовательно, по срокам пострадиационного восстановления клетки крови располагаются в следующем убывающем порядке: эритроциты, нейтрофилы, тромбоциты и лимфоциты. Такая последовательность совпадает с разрозненными данными о последовательности восстановления клеток крови у людей, перенесших лучевую болезнь.
Далее следует еще отметить, что ни у одной из наблюдавшихся собак не удалось и через 3 года после облучения выявить нормализации лимфопоэза. В то же время показатели красной крови первыми достигали нормальных величин. Эта нормализация сопровождалась заметным увеличением числа ретикулоцитов, т. е. увеличенным выбросом из костного мозга в периферическую кровь недозревших эритроцитов. Это, а также сохранившийся сдвиг отношения бластных клеток белого и красного рядов в сторону красного свидетельствует о том, что нормализация содержания эритроцитов и гемоглобина в периферической крови происходит за счет напряжения эритропоэза.
По мнению многих специалистов, изменения красной крови в большинстве случаев прогностически более важны, чем белой крови. В организме отдается предпочтение восстановлению той крайне необходимой для организма дыхательной функции, какую выполняет красная кровь. Это позволило считать, что без истинной нормализации эритропоэза в преобладающем большинстве случаев трудно ожидать стабильного восстановления других ростков кроветворения и их функций, в частности иммунитета.
Однако представления о состоянии эритропоэза, полученные по данным общепринятых исследований цитологического состава крови и костного мозга, не всегда могут правильно отражать состояние кроветворения. Миелограммы в ряде случаев не позволяют судить о функциональной активности всей гемопоэтической ткани в силу возможной очаговости изменений в ней и в случаях изменения плацдарма кроветворения (т. е. массы кроветворной ткани). По данным пунктатов костного мозга трудно судить о количественных изменениях в скорости кроветворения, особенно когда нет изменений в соотношениях отдельных стадий созревания клеток. Поэтому суммарную продуктивность эритропоэза определяли по включению 59Fe в эритроциты. Меченое железо включалось на стадии гемоглобинизации эритроцитов. Расчеты меченого железа, поглощенного эритроцитами 1 мл крови, 1 млн. эритроцитов или 1 г гемоглобина, давали идентичные результаты. В этих условиях радиоактивность циркулирующих эритроцитов характеризовала продукцию их всей массой кроветворной ткани и поступление их в периферическую кровь. Примененная методика исключала влияние возможных часовых, суточных и ситуационных колебаний в течение указанных процессов па конечный результат, поскольку определялось интегральное поглощение 59Fe за четырехсуточный период.
Изучены суммарная интенсивность эритропоэза у собак через 1; 2,5; 12 и 42 мес. после облучения в дозе 4 Гр и показатели у других групп собак, подвергавшихся облучению в разных дозах. Выявлены наиболее характерные результаты для собак, у которых после перенесения острого лучевого поражения цитологические показатели крови не имели сколько-нибудь выраженных изменений или наблюдались умеренные изменения со стороны белой крови. После однократных облучений с конца первого месяца после лучевого воздействия и далее в различные сроки до 3,5 лет у многих, а в определенных условиях у большинства, собак отмечался повышенный темп эритропоэза при нормальном или несколько пониженном уровне лейкоцитов и значительно реже эритроцитов и гемоглобина. Часто, но не всегда обнаруживался ретикулоцитоз. Нередко наибольшее включение 59Fe в эритроциты наблюдалось у тех животных, у которых была более выраженная эритропения. Особенно значительное усиление темпа кроветворения (в 2 раза и более) отмечалось после облучения в дозах 2,0—3,5 Гр (дозы, близкие или несколько превышающие максимальные несмертельные, но вызывающие клинически выраженную лучевую болезнь).
Повышенный темп эритропоэза при нормальной периферической крови можно рассматривать как подтверждение того, что восстановление и нормализация системы красной крови, связанной с функциями, жизненно необходимыми для существования организма, происходят в первую очередь. Отсутствие изменений в цитологическом составе крови не должно служить доказательством истинного благополучия гемоноэза. Напряжение эритропоэза, отмеченное в течение весьма длительного времени после лучевого воздействия, может быть одной из причин задержки в восстановлении числа лейкоцитов (лимфоцитов) или тромбоцитов у перенесших лучевое поражение или отсутствия устойчивой нормализации в их уровне. Другими словами, без истинной нормализации эритропоэза не следует ожидать стабильного восстановления других ростков кроветворения и выполняемых ими функций, в первую очередь иммунитета. Отмечаемые в течение длительных периодов пострадиационные лейкопенические состояния с этих позиции следует, по-видимому, рассматривать как обусловленные преимущественной пролиферацией эритробластических элементов, что вызывалось необходимостью первоочередного восстановления и поддержания функций красной крови в пределах физиологической нормы.
Выявленные изменения в интенсивности гемопоэза нельзя однозначно связывать с глубиной радиационного поражения системы крови. Повышение темпа эритропоэза можно рассматривать как компенсаторно-приспособительную реакцию, направленную на поддержание функций красной крови в пределах физиологической нормы. Степень выраженности этой реакции определяется двумя главными факторами: необходимостью и возможностью ее развития. Необходимость повышения темпа эритропоэза возникает при ускоренной убыли эритроцитов из сосудистого русла. Выраженность этой реакции зависит от пределов физиологических возможностей регенеративного аппарата эритропоэза и глубины его радиационного поражения.
Продолжительность жизни эритроцитов в сосудистом русле у изучавшихся собак, определявшаяся с помощью метки собственных эритроцитов 51Сг, оказалась сокращенной. У одной опытной и у одной контрольной здоровой собаки одновременно определяли и интенсивность эритропоэза, и эритрофагоцитарную функцию селезенки через 190 суток после окончания многократного лучевого воздействия, используя 51Сг в качестве метки. С указанной целью через 2 недели после введения в организм меченных изотопом собственных эритроцитов извлекали селезенку и готовили срезы толщиной около 0,5 см. Срезы промывали физиологическим раствором для удаления крови, после чего определяли их радиоактивность, средняя величина которой выражалась в процентах от радиоактивности селезенки контрольной собаки. У облученной собаки радиоактивность была в 1,5 раза больше, что свидетельствовало об усилении фагоцитоза эритроцитов клетками селезенки.
Подробное описание состояния указанных групп собак дано в наших монографиях [Акоев, 1970; Акоев и др., 1981]. Средние данные по 12 собакам через 190 сут после окончания многократного облучения: внешнее клиническое благополучие, морфологический состав крови у облученных собак не выходил за пределы нормы. Однако отмечалось относительное снижение числа лейкоцитов (на 2,9 тыс. клеток в 1 мм3 крови) и повышение содержания ретикулоцитов (3,1% против 1,2% в контрольной группе). Это давало повод предполагать скрытое напряжение эритропоэза. И действительно, оно подтвердилось исследованиями с меченым железом. Суммарное поступление эритроцитов в русло крови оказалось в 1,75 раза выше нормального, что, в свою очередь, могло служить косвенным доказательством ускоренного разрушения эритроцитов.
Возможность очень длительного сохранения сокращенной продолжительности жизни эритроцитов у человека после радиационных воздействий была показана Хубером и Ширакурой на примере исследований, проводившихся не менее чем через 6 лет после успешной лучевой терапии 25 женщин по поводу рака матки и обнаруживших у них уменьшенную продолжительность пребывания эритроцитов в сосудистом русле.
Изложенные исследования, выполненные нами на собаках, демонстрируют определенную последовательность включения и выключения разных механизмов усиления продуктивной способности гемопоэза, необходимых для поддержания в первую очередь функции красной крови. Из анализа представленных данных можно заключить, что примат производства клеток красного ростка существует тогда, когда имеется значимая для организма недостаточность количества эритроцитов и гемоглобина. Первым мобилизуемым резервом является расширение плацдарма кроветворения, т. е. увеличение массы гемопоэтической ткани. Этот резерв — расширенный плацдарм кроветворения — используется организмом наиболее длительно. У собак в этот период «костный мозг» грудины и энифимзов на секции становится красным, т. е. происходит замещение его эритропоэтической тканью.
Вторым возможным мобилизуемым резервом является ускорение прохождения клетками делящегося и делящегося-созревающего пулов. Наконец, позже всех и на самое короткое время (на месяц-полтора) может увеличиваться митотический индекс эритробластов, непосредственно отражающий в этих условиях сокращение их генерационного цикла.
Основные наблюдения за состоянием кроветворения у собак в отдаленный пострадиационный период (изменения периферической крови и интенсивность эритропоэза) были подтверждены нами и в экспериментах на крысах.
Возможность сокращения продолжительности генеративного цикла бластных клеток белого и красного ряда была доказана А.В. Илюхиным с соавторами при длительном хроническом воздействии радиации на собак. После введения фенилгидразина или массивного кровопускания также обнаружено сокращение генерационного времени делящихся эритробластов, которое в основном происходило за счет стадии Gt.
Была подробно изучена цитокинетика кроветворения у собак на протяжении трехлетнего слабого хронического воздействия радиации. Собак в течение трех лет непрерывно (кроме времени на кормление и обследование) облучали: суммарные дозы от 0,63 до 5,70 Гр.
В периферической крови на 4—12-м месяце облучения отмечали ретикулоцитоз, сокращение времени пребывания эритроцитов в крови (на 12—24-м месяце) при сохранении числа эритроцитов и содержании гемоглобина в пределах контрольных значений (табл. 2). Это могло указывать па скрытое напряжение эритропоэза для компенсации сокращения продолжительности жизни эритроцитов и поддержания увеличенной скорости обновления эритроцитов.
Оценка костномозговой продукции эритроцитов показала статистически значимое ее увеличение, сохранявшееся в течение всего второго года облучения. Тем не менее качество эритроцитов (насыщенность гемоглобином, кривые Прайс—Джонса) не ухудшалось. К концу третьего года все показатели были в пределах нормы.
Со стороны белой крови отмечали стойкое снижение лейкоцитов и нейтрофилов па протяжении всего третьего года облучения. Продолжительность пребывания грану
Таблица 2. Сроки (в месяцах) выявления статистически достоверных отклонений в цитокинетических пикалагелях эритропоэза и лейкогранулопоэза у собак на протяжении трехлетнего хронического радиационного воздействия в малых дозах (наша обработка данных А.В. Илюхина и др.)
| Показатель | Эритропоэз | Лейкогранулопоэз | ||
| Увеличение | ||||
| Снижение | Увеличение | Снижение | ||
| Число клеток в периферической крови |
4-12 (ре- тикуло- циты) |
- | 24-36 | |
| Период циркуляции клеток в крови | 12-24 | 32 | ||
| Скорость обновления клеток в крови | 24 | 32 | ||
| Костно-мозговая продукция зрелых клеток | 12-24 | 36 | ||
| Качество зрелых клеток | 12-36 | |||
| Число клеток костного мозга общее | 28-36 | 24-36 | ||
| Скорость обновления клеток костного мозга | 28 | 28-26 (до 2 раз) | ||
| То же для отдельных властных форм | 24-32 | |||
| Длительность генерационного цикла | 28 (Gi) |
28-36 (G..S, G2) |
||
| Костно-мозговой резерв | 5-36 |
Осмотическая стойкость лейкоцитов была сниженной на протяжении последних 2,5 лет наблюдений. В отличие от показателей красной крови, которые нормализовались на протяжении третьего года облучения, в белой крови продолжала к этому сроку нарастать недостаточность миелопоэза. Изменения показателей красной крови в условиях слабого хронического воздействия были менее выраженными и восстанавливались быстрее, явно отдавая приоритет восстановлению эритропоэза. Авторы пишут о возможном дефиците стволовых клеток и их направленной дифференцировке в эритроидном направлении.
Эти заключения авторы подтверждают анализом и непосредственных изменений костного мозга указанных собак. Средняя продолжительность генерационного цикла миелоидных элементов изменялась существенно — почти в 2 раза. В отличие от цикла эритроидных элементов (где статистически достоверно сокращалась лишь стадия Gi) здесь укорачивались все стадии интерфазы клеточного цикла. С помощью пирогеналовой пробы обнаружили снижение костномозгового резерва гранулоцитов у большинства собак в большинстве сроков исследовании, в то же время не выявлено уменьшение резерва эритроидного ростка.
Таким образом, обобщая данные о состоянии кроветворения в период длительного слабого хронического воздействия радиации п в период отдаленного пострадиационного восстановления после острых и многократных облучений, можно заключить, что наиболее характерным для гемопоэза было явное или скрытое напряжение эритропоэза, в значительной мере определявшее состояние периферической крови и клеточность костного мозга. Значительную роль при этом играет сокращенная продолжительность жизни эритроцитов. Основной вклад в пополнение клеточности костного мозга после экстремальных воздействий вносят процессы пролиферации и дифференцировки морфологически не различимых стволовых клеток костного мозга. Они обладают чрезвычайными потенциями к пролиферации и имеют другие возможности усиления продуктивности отдельных ростков кроветворения.
2. Регуляция размножения и дифференцировки кроветворных клеток
В связи с успехами исследования процессов кроветворения и функциональных возможностей стволовых клеток при использовании метода селезеночных колоний и культур на полутвердых средах и других методов представления о регуляции путей их пролиферации и дифференцировки существенно изменились.
Экспериментально подтвердилось существование единой полипотентной стволовой клетки, способной к дифференцировкам по всем направлениям гемопоэза, включая лимфопоэз (рис. 3). Казалось бы, это непосредственно доказывает правильность исходного положения о конкуренции различных ростков кроветворения за направление дифференцировки стволовой клетки в соответствии с потребностями организма. Однако вопрос оказался значительно сложнее.
Наряду с исходной полипотентной стволовой кроветворной клеткой могут существовать и обладающие стволовыми свойствами предшественники, коммутированные (т. е. ограниченные к выбору направления дифференцировки) предшественники двух типов — миелопоэза и лимфопоэза. Это первое и главное разделение направлений возможной дифференцировки единой полипотентной стволовой клетки. Как осуществляется регуляция выбора дифференцировки — в сторону миелопоэза или лимфопоэза, не очень ясно.
Стволовые клетки, коммутированные в сторону лимфопоэза, дифференцируются сначала в общих, а затеи в раздельных предшественников Т- и В-лимфоцитов, из которых через ряд промежуточных стадий образуются Т-лимфоциты (приобретая определенные клеточные рецепторы) и В-лимфоциты (приобретая рецепторы к комплементу, к фрагментам и комплексам иммуноглобулинов и комплексам антиген—антитело). Кроме того, из них образуются плазматические клетки. Лимфопоэз наименее изучен. До сих пор неясно, имеются ли в лимфоцитарном ряду клетки, не способные к пролиферации. Среди лимфоидных клеток трудно определить принадлежность их к пролиферирующему, созревающему или функциональному пулам, так как возможны взаимные перекрытия.
Стволовые клетки, коммутированные в сторону миелопоэза, в дальнейшем через поэтинчувствительные предшественники разделяются на три ростка: эритроцитарный, гранулоцито-макрофагальный и тромбоцитарный.
В эритроцитарном направлении выявлено несколько промежуточных клеток (в том числе и бурстообразующая клетка) между общим предшественником миелопоэза и эритропоэтинчувствительными клетками. Последующие морфологически различаемые клетки относятся к про-эритробласту, трем видам эритробластов, ретикулоциту и зрелому эритроциту. Обычно происходит 5—6 митозов в морфологически различимых клетках, и каждый эритробласт дает 30—60 эритроцитов. Эритробласты теряют способность к делению, как только в клетке накопится 27 нг. гемоглобина. В случае замедления синтеза гемоглобина увеличивается число делений эритробластов. Из одной полипотентной стволовой клетки, начавшей дифференцировку в сторону миелопоэза, может образоваться около 1 млн эритропоэтинчувствительных клеток и около 0,1 млн. клеток — предшественников гранулоцитов и макрофагов,
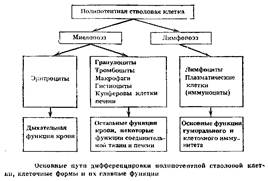
Развитие гранулоцито-макрофагального направления миелопоэза проходит на стадии образования клеток, дающих колонии на полутвердых средах (KOEJ, кластерообразующих клеток и предшественников гранулоцитов и моноцитов-макрофагов. Последующие клетки морфологически различимы. Миелобласт нейтрофильный далее дает клетки гранулоцитарного ряда (эсзинофил), нейтрофильного ряда (нейтрофил) и базофильного ряда (базофил тучная клетка).
Монобласт дает моноциты, макрофаги, гистиоциты, купферовы клетки печени и остеокласты. Клетки моноцитопоэтического ряда объединены в систему фагоцитирующих мононуклеаров как способные к фагоцитозу и осуществляющие в норме функции клеточного фагоцитоза. Ранее входившие вместе с указанными клетками в ретикуло-эндотелиальную систему ретикулярные клетки и эндотелиальные клетки исключены из системы фагоцитирующих мононуклеаров как являющиеся лишь факультативными фагоцитами, не имеющими рецепторов для иммуноглобулинов и не способными к иммунному фагоцитозу. Клетки же системы фагоцитирующих мононуклеаров в процессе дифференцировки приобретают рецепторы для иммуноглобулинов и комплемента и становятся способными к активному иммунному фагоцитозу.
Тромбопоэтинчувствительные клетки формируют морфологически различимые клетки: мегакариобласт, проме-гакариоцит и тромбоциты. Для предшественников характерны 4—5 эндомитозов.
Таким образом, по современным данным, клетки костного мозга, способные к делению, можно условно разделить на морфологически различимые клетки, как правило, способные к очень ограниченному числу делений (4— 8 делений), и на морфологически неразличимые клетки, выявляемые лишь по способности их давать колонии в селезенке облученных мышей или пролиферировать на полутвердых средах (и другими методами), способные к чрезвычайно активной пролиферации. Число делений, которые может совершить полипотентная стволовая клетка, на порядок больше, чем для морфологически различимых клеток. В норме их в костном мозге лишь около 0,5% от числа всех клеток. Однако достаточно сохраниться 0,1% стволовых клеток после сильного радиационного воздействия, чтобы обеспечить восстановление нормального пула стволовых клеток и резкое повышение дифференцировки клеток в последующих отделах.
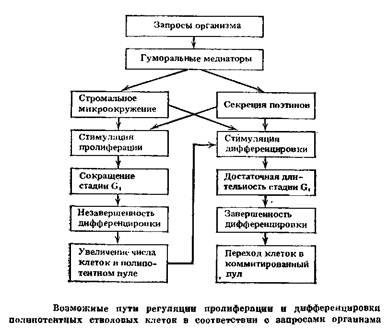
При нормальном кроветворении абсолютное большинство стволовых клеток находится вне клеточного цикла (период Go или длительный период Gi). Стабильное поддержание величины отдела стволовых клеток обеспечивается небольшой долей клеток, вступивших в митотический цикл. В случае уменьшения отдела стволовых клеток в пролиферацию может вовлекаться вся популяция сохранившихся стволовых клеток. При этом время удвоения популяции стволовых клеток оказывается очень коротким, порядка 15—20 ч. По данным ряда авторов, укорочение или удлинение стадии Gi стволовых клеток соответственно сопровождается пролиферацией (т. е. оставлением в пуле стволовых клеток) или дифференцировкой (т. е. уходом в последующие отделы).
Следовательно, основной вклад в восполнение дефицита клеточности костного мозга вносит пролиферация морфологически неразличимых клеток и потому исследование регуляции их численности и регуляции выбора направления дифференцировки является чрезвычайно важным. К сожалению, в этих вопросах еще очень много неясного.
Как решается для стволовых клеток первый выбор направления дифференцировки (в сторону лимфопоэза или миелопоэза), тоже неясно. По многочисленным данным, в опытах на собаках и обезьянах аллогенная кроветворная ткань вызывала появление огромного количества лимфоцитов, специфически иммунных против реципиента, при полной или частичной блокаде остального кроветворения. Следовательно, возможно переключение части стволовых клеток с эритроидного на лимфоидный путь дифференцировки. Характер дифференцировки кроветворной ткани в условиях трансплантации указывает на конкуренцию миелоидных и лимфоидных клеток за общий предшественник. У полицитемических радиохимер, у которых снижен эритропоэз, ускоряется восстановление лимфопоэза. Приведенные нами данные о существенной задержке пострадиационного восстановления числа лимфоцитов в крови человека в период, когда эритропоэз напряжен, также свидетельствуют о конкурентных отношениях ростков кроветворения. Об этом непосредственно говорит и проведенное нами изучение соотношений в морфологически различаемых клетках разных ростков костного мозга у облучавшихся собак.
Тем не менее, некоторыми учеными предполагается, что направление дифференцировки стволовых клеток или закреплено генетически постоянно, или осуществляется стохастически, в обоих случаях — независимо от запросов организма. Возможным доказательством этого может служить одно и то же распределение маркерных клеток по разным росткам гемопоэза при изменении общих запросов организма.
В связи с изложенным, вероятно, более правильно считать существование конкурентных отношений между ростками кроветворения за преимущественную продукцию конечных зрелых форм клеток, а не примат дифференцировки полипотентной стволовой клетки в тот или иной росток гемопоэза. При этом влияние дальнодействующей для гемопоэза регуляции с целью реакций па запросы организма в экстремальных ситуациях может осуществляться не только через поэтиподобные регуляторы, но и через микроокружение в локальных участках гемопоэтических тканей, которое может зависеть также от запросов организма. Эритропоэтин для красного ростка и колониестимулирующий фактор для гранулоцитарного ростка стимулируют пролиферацию клеток и укорачивают средний период генерации клеток. При этом удлинение пли укорочение стадии Gi клеточного цикла может определять уход клеток в пролиферацию или дифференцировку (рис. 4). В случае вступления клетки в деление сокращается генерационный цикл и клетка не успевает осуществить дифференцировку. Пролиферация прекращается, как только весь объем микроокружения окажется занятым стволовыми клетками.
Таким образом, исследования реакции отдела стволовых клеток на экстремальные воздействия и пострадиационные изменения гемопоэза позволили показать ряд общих закономерностей в реакции этой системы на экстремальные воздействия п в процессах последующего восстановления.
Влияние запросов организма на гемопоэз осуществляется дальнодействующей регуляцией посредством гуморальных факторов. Поэтому следует кратко рассмотреть отдельные радиационные изменения со стороны эндокринной и других систем организма.
3. Другие проявления отдаленных радиационных последствий в предпатологический период
Обобщение и анализ остаточных и отдаленных последствий воздействия радиации проводились неоднократно. Наряду с изменениями в системе крови, изложенными выше, отмечались последствия и в других системах организма.
К ним в первую очередь необходимо отнести нарушения иммунологической реактивности, проявляемые наиболее четко в снижении устойчивости к инфекционным агентам. Так, устойчивость к живому возбудителю газовой гангрены и его токсину в экспериментах была снижена у мышей через 190 сут., а у крыс — через 150 сут. после воздействия радиации.
Имеются также данные, что для полного восстановления иммунобиологической реактивности у облученных грызунов необходимы чрезвычайно длительные сроки, составляющие значительную часть всей нормальной продолжительности жизни животных. Это хорошо соответствует изложенным выше данным о длительной задержке восстановления лимфоидной и гранулоцитарной части кроветворения после облучения.
Цикл исследований был выполнен нами на собаках через 190 сут. после окончания многократного воздействия (осуществлявшегося в 2—4 приема на протяжении 2,5—14 мес.) при полном клиническом благополучии животных и гематологических показателях в пределах видовой нормы к моменту исследования. О скрытой неполноценности восстановления крови можно было только предполагать: число лейкоцитов несколько ниже, чем в контроле, фагоцитарная активность лейкоцитов (число фагоцитирующих клеток) и фагоцитарный индекс (количество микробов на один фагоцит) недостоверно выше, а абсолютный фагоцитарный показатель (число введенных в кровь клеток золотистого стафилококка, фагоцитированных лейкоцитами 1 мм3 крови) был на 25% ниже контрольного уровня. Кроме того, у одной из собак при некотором снижении числа эритроцитов обнаружена в 1,75 раза более высокая суммарная продуктивность эритропоэза. Следовательно, отмечалась типичная картина скрытого напряжения гемопоэза, прикрытого стандартными показателями периферической крови в пределах видовой нормы или несущественно отличаясь от нее.
Эффективной проверкой состояния систем организма собак спустя 190 сут. после окончания многократного воздействия радиации являлась реакция на травматическое повреждение и скорость посттравматической регенерации. С этой целью на спину собак в области между лопатками наносили кожную рану путем отсечения кожного лоскута. Рану антисептической обработке не подвергали. Оценивали период времени, необходимый для уменьшения площади раневой поверхности наполовину. Период полурепарации раны у ранее облучавшихся собак был больше, чем у контрольных животных, имея тенденцию к возрастанию с увеличением суммарной дозы облучения. Количество лейкоцитов, участвовавших в фагоцитозе введенного в кровь золотистого стафилококка, у некоторых животных резко увеличилось, в 1,5—2,5 раза превысив исходный уровень. У ранее облучавшихся собак, наоборот, отмечено уменьшение почти в 2 раза исходного и контрольного уровней фагоцитарной активности лейкоцитов. Еще большие различия выявлены в абсолютном фагоцитарном показателе: например, на 21-е сутки после ранения он был равен у контрольных животных 29,8 тыс., а у облученных собак — 5,5 тыс. микробных тел, фагоцитированных лейкоцитами 1 мм3 крови.
В экспериментах на облученных мышах с закрытым переломом костей голени или стандартной кожной раной также обнаружено замедление посттравматической регенерации, в том числе и в случае, когда животным наносились травмы через месяц после четырехкратного с месячным интервалом облучения в несмертельных дозах. Относительная величина удельного замедления репарации при заживлении костной ткани была больше, чем кожной.
Сниженную устойчивость собак к повторному воздействию радиации наблюдали через 10 мес. после окончания многократного воздействия радиации.
Известно, что изменения со стороны центральной нервной системы могут отмечаться у перенесших лучевое воздействие в течение длительных периодов времени. Более того, со временем они у ряда лиц могут быть более выраженными. У некоторых лиц, подвергавшихся субтотальной рентгенотерапии, явления астенизации прогрессивно нарастали в течение 3—4 лет и в дальнейшем па протяжении 5—7 лет и более перерастали в симптомы органического поражения нервной системы.
К последствиям, для которых характерен длительный латентный период после воздействия радиации, относят дегенеративно-дистрофические и склеротические изменения в различных органах и системах. Атрофические, склеротические и язвенные изменения слизистой желудочно-кишечного тракта, циррозы печени, невросклерозы и др. В.Н. Стрельцова и Ю.И. Москалев относят к неопухолевым формам поздних эффектов действия радиации. О развитии в поздние сроки радиационных функциональных и морфологических изменений эндокринных желез сообщалось в ряде работ. В.В. Шиходыров и соавторы отмечают некоторые различия в реакции ткани на грубое структурное поражение (обычно в критических органах) и на менее выраженное поражение (обычно в некритических органах). В обоих случаях изменения не зависят от характера поврежденного агента.
В первом случае начальным звеном процесса служат некробиотические изменения ткани. Наряду с процессом дистрофии развиваются явления склероза. Разрастание соединительной ткани оценивается вторичным, заместительным процессом в ответ на уменьшение числа клеток паренхимы органа. Определенное значение в развитии упомянутых нарушений имеют изменения кровеносных сосудов.
Во втором случае не возникают первоначальные грубые структурные изменения паренхимы органа. В течение длительного времени отсутствуют какие-либо морфологические повреждения. Происходят постепенное развитие патологического процесса, уменьшение клеточности паренхимы органа и нарастание атрофии и склероза. Изложенные процессы в целом имеют ту же направленность, что и при старении. У облученных животных эти процессы происходят более ускоренно. Так, однотипные изменения в центральной нервной системе наблюдаются у интактных собак в 7—12 лет и у облучавшихся — в 5—7 лет. Важно отметить, что дистрофические изменения и нарушение процессов физиологической регенерации часто могут сопровождаться развитием очагов узловатой гиперплазии.
Отмечается, что в условиях длительно текущих и выраженных дистрофических нарушений во внутренних органах возникает повышенная нагрузка на органы эндокринной системы, связанная с возможным перенапряжением и истощением их. В менее демонстративной форме такое влияние на старение оказывают и другие факторы (инфекционные заболевания, интоксикация, нарушения питания и др.).
Для пострадиационных изменении иммунитета наиболее характерны сочетания иммунодефицитного и ауто-агрессивного состояний, недостаточность механизмов неспецифической и специфической иммунной защиты организма. Общая иммунологическая реактивность после введения в организм ряда радионуклидов изменялась при таких низких суммарных дозах, как 0,3 Гр за 30 сут. для стронция и 0,6 Гр за 30 сут. для цезия. Фагоцитарные реакции изменялись примерно при таких же низких дозах воздействия. Еще более низкие дозы необходимы, чтобы началась атрофия лимфоидной ткани и плазматизация селезенки (0,10—0,35 Гр за год воздействия). Примерно при таких же суммарных поглощенных дозах происходили изменения в образовании аутоантител (увеличение их). Изложенное свидетельствует о чрезвычайной чувствительности отдельных реакций иммунитета к внешним воздействиям. Изменение соотношения числа Т- и В-лимфоцитов также отмечали при суммарных поглощенных дозах порядка 0,1 Гр. Нелетальное воздействие радиации может вызывать ряд аутоиммунных и аллергических расстройств (данные В.М. Шубина).
Различают первичную (генетически обусловленную) и вторичную (индуцированную) иммунологическую недостаточность. Последняя может развиваться как после воздействия ионизирующих излучений, так и после приема иммунодепрессантов, а также при старении и некоторых заболеваниях. Все эти средства и состояния неспецифически угнетают и клеточный и гуморальный иммунитет.
Особенностью развития аутоиммунной реакции при действии ионизирующей радиации и других воздействий в отличие от иммунного ответа на чужеродные антигены является ее формирование не в сторону развития невосприимчивости, а в сторону сенсибилизации, т. е. повышения чувствительности к тканевым продуктам распада. Образование аутоантител происходит не только при действии излучения, но и при иммунизации микробными антигенами. Иммуногенез после облучения угнетен по отношению к чужеродным агентам, а по отношению к собственным тканям, наоборот, активно развивается.
Известно, что в тканях (особенно в селезенке) здорового организма, не имеющего контакта с инфекцией или предупреждающими воздействиями, всегда можно обнаружить нормальные клетки, образующие аутогемолизины. В крови их число доходит до 3% ядросодержащих клеток, а в паренхиматозных органах — в пределах 5— 10% Они выполняют важную физиологическую роль по удалению старых клеток и обезвреживанию продуктов их распада. Однако под влиянием радиации, других повреждающих факторов и старения эта функция перерастает в аутоагрессивную и становится сама повреждающим фактором.
Многие клинические формы и патологические состояния не являются специфическими для радиационных последствий. Они известны и для возрастной патологии. В связи с этим широко обсуждаются и вопросы ускорения и увеличения частоты возрастных изменений в результате действия радиации.
Особый интерес вызывает определенная последовательность изменений в эндокринной системе, играющей важную роль в регуляторных механизмах взаимодействия систем организма. Анализ временной последовательности изменений в эндокринной системе и становления клинических форм отдаленных радиационных последствий позволил нам прийти к выводу о зависимости многих проявлений радиационных последствий от недостаточности эндокринных органов.
По данным Е.Н. Антипенко, изучавшего функциональное состояние и структуру щитовидной железы у собак, через 3,5 мес. после лучевого воздействия в дозе 4 Гр йодфиксирующая функция еще не отличалась от нормы. Однако при гистологическом исследовании были обнаружены участки с пониженной и повышенной тиреоидной активностью. Через 6—13 мес. после облучения деятельность железы усиливалась, о чем свидетельствовали кривые накопления радиойода в железе, содержание органически связанного радиойода в плазме крови и строение железы. Характерно, что явлений тиреотоксикоза не было. Однако в этот период продолжали нарастать со временем частота, а также выраженность дистрофических и некробиологических изменений. Масса железы снижалась. Выше приводились данные о состоянии кроветворения у собак в эти же временные отрезки: скрытое напряжение эритропоэза при нормальной, но нестабильной картине крови или при незначительном снижении числа лейкоцитов и лимфоцитов или эритроцитов при увеличении числа ретикулоцитов, костный мозг гиперпластичен по красному ростку, продолжительность жизни эритроцитов сокращена.
Через 5 мес. после облучения крыс в дозе 6 Гр нами проводился комбинированный опыт. Величина максимального накопления радиойода в железе крыс была в 1,5 раза выше, чем в контроле. Скорость накопления u выведения его также оказалась выше. Относительная масса железа на 100 г массы тела была ниже, чем у контрольных крыс. Усиленная йодфиксирующая способность железы, относительно невысокое содержание в ней коллоида и его повышенная резорбция, а также полнокровие железы свидетельствовали о значительном усиление гормонообразования и особенно гормоновыделения без явлений тиреотоксикоза.
Одновременно у части крыс анализировали состояние кроветворения. У облученных животных отмечали тенденцию к снижению числа эритроцитов и гемоглобина, повышение ретикулоцитов в крови и увеличение суммарной продуктивности эритроцитов по данным включения радиоактивного железа. Эта тенденция к напряжению эритропоэза оказалась менее выраженной, чем у более молодых крыс (опыт ставили на крысах в возрасте 1,5 лет, у которых можно было ожидать более яркого проявления гормональной терапии в связи с возможным начинающимся возрастным угасанием эндокринных функций). У отдельных животных отмечалось, наоборот, значительное снижение скорости включения меченого железа и уменьшение количества эритроцитов.
У другой группы крыс определяли устойчивость к повторному облучению. Полулетальная доза повторного облучения, которое проводили через 5 мес. после первого воздействия радиации, оказалась на 1,47 Гр меньше, чем в контроле. Устойчивость ранее облучавшихся крыс к токсину возбудителя газовой гангрены в дозе, близкой к минимальной абсолютно смертельной, оказалась также сниженной. Среднее время выживаемости сократилось по сравнению с контролем.
Учитывая, что усиленная функция щитовидной железы у облучавшихся животных в указанных опытах не сопровождалась явлениями токсикоза и могла быть расценена как компенсаторная в связи с возросшими потребностями организма, в последнем опыте на крысах нами проверялась эффективность введения в организм гормонов щитовидной железы (тиреоидин, который давали с пищей ежедневно на протяжении четвертого и пятого месяцев после облучения, или диэтилстильбестрол, который добавляли в пищу в те же сроки).
У ранее облучавшихся крыс, получавших гормоны щитовидной железы, отмечали значительное снижение функций щитовидной железы (йодфиксирующая функция резко снижена, относительная масса железы ниже контрольной, железы образованы крупными фолликулами с большим количеством гомогенного коллоида); повышение устойчивости к повторному воздействию радиации, которое, однако, не достигло контроля; повышение устойчивости крыс к токсину возбудителя газовой гангрены (по не достигло контрольного уровня); снятие напряжения эритропоэза (число эритроцитов и содержание гемоглобина повысились, число ретикулоцитов снизилось, продукция эритроцитов уменьшилась до контрольного уровня).
В этом опыте введение тиреоидина ранее облучавшимся крысам оказало благотворное действие на устойчивость организма к внешним воздействия, на щитовидную железу, на гемопоэз, несмотря на усиление гормонообразовательной и инкреторной функций железы у не получавших тиреоидина крыс. Это доказало, что в организме в этот период не хватало гормонов щитовидной железы, несмотря на усиленное поступление их из железы, а усиление функций железы носило компенсаторно-приспособительный характер в ответ на повышенное расходование их в облученном организме. Дополнительное введение гормонов в организм по этой причине не вызывало токсикоза, а оказывало благоприятное действие. Гормональный дефицит и указанные отдаленные последствия оказались в определенной связи.
Другая группа крыс получала половой гормон. У крыс этой группы йодфиксирующая функция щитовидной железы была чрезмерно увеличена. Относительная масса железы увеличилась, фолликулы были образованы гипертрофированными клетками, запас коллоида практически отсутствовал. Это чрезмерное напряжение гормонообразовательной и инкреторной функций щитовидной железы могло быть обусловлено значительным усилением тиреотропной функции гипофиза за счет снижения гонадотропной в связи с насыщением организма половым гормоном. Устойчивость к повторному воздействию радиации повысилась. 'Введение в организм полового гормона привело не только к уменьшению недостатка в нем (крысы были к этому времени в возрасте двух лет, т. е. нерепродуктивного возраста), но и через дальнейшее усиление функций щитовидной железы к снижению дефицита в гормоне последней. У животных под влиянием введения полового гормона наблюдали увеличение содержания гемоглобина при сохранении числа эритроцитов на прежнем уровне, увеличение числа ретикулоцитов и тенденцию к дальнейшему увеличению суммарной продукции эритроцитов костным мозгом. Таким образом, если введение тиреоидина в организм крыс через 5 мес. после облучения способствовало ликвидации напряжения эритропоэза и гиперфункционального состояния щитовидной железы, предотвращая возможность их функционального истощения, то введение полового гормона, наоборот, вызывало дальнейшее функциональное напряжение гемопоэза и щитовидной железы, усугубляло возможность истощения функций и перехода их в патологическую сверхкомпенсацию.
В наших исследованиях на собаках, которым наносили через 190 сут. после окончания многократного облучения хирургическую травму, также испытывали действие предварительного 15-суточного введения тиреоидина с пищей или преднизолона (препарат коры надпочечников). В группе собак, получавших тиреоидин перед нанесением травмы, количество эритроцитов и гемоглобина осталось таким же, как и в группе облученных, но не получавших гормонального препарата. Число ретикулоцитов снизилось до нормального уровня. Продукция эритроцитов, определявшаяся по включению меченого железа, снизилась. Число лейкоцитов в 1 мм3 крови поднялось. Резко возросли показатели фагоцитарной активности лейкоцитов: число фагоцитирующих золотистый стафилококк клеток, количество поглощенных микробов на один фагоцит и число микробов, фагоцитированных лейкоцитами 1 мм3 крови, увеличились в 2—2,5 раза и более по сравнению с облученным контролем и здоровыми животными.
В группе собак, получавших преднизолон, несколько увеличилось число эритроцитов и лейкоцитов и уменьшилась продукция костным мозгом эритроцитов. Содержание гемоглобина, число ретикулоцитов не изменились. Показатели фагоцитарной активности лейкоцитов крови также существенно были более высокими.
Однако после ранения собак выявилась неполноценность функции фагоцитоза. Высокая фагоцитарная активность лейкоцитов до нанесения раны, вызванная введением в облученный организм преднизолона, оказалась менее полноценной, чем более умеренное участие лейкоцитов в фагоцитозе микробных тел у здорового организма. Это обусловлено тем, что положительное влияние эндокринных препаратов на фагоцитарный индекс, т. е. на количество микробов, поглощаемых каждым лейкоцитом, на фоне сниженного числа лейкоцитов в крови оказалось недостаточным, чтобы абсолютный фагоцитарный показатель после ранения был таким же, как у здоровых животных. Тем не менее, гормонотерапия все же позволила весьма существенно поднять этот показатель (например, на 21-й день после ранения с 5,5 до 12,2 тыс. и до 20,2 тыс. микробов, фагоцитированных лейкоцитами 1 мм3 крови соответственно в группах, получавших тиреоидин и преднизолон).
В другом опыте, проводившемся на живых мышах через 5 мес. после воздействия радиации в дозе 5,75 Гр, вызвавшем гибель У3 животных в течение 4 мес., также определяли функциональное состояние щитовидной железы. У этих животных поглощение радиойода щитовидной железой составляло 60% от введенного количества изотопа, что существенно ниже по сравнению с контролем. Активность связанного с белком йода также была ниже контроля. Все это позволило предполагать наступающее истощение функций щитовидной железы. Морфологические исследования подтвердили это заключение.
Следует сказать, что смену повышенной деятельности щитовидной железы гипофункциональным состоянием после радиационного воздействия отмечали многие исследователи. Так, у крыс через 40 сут. после однократного введения 210Ро наблюдали морфологические признаки гиперфункцнонального состояния щитовидной железы, а через 70—100 сут. —явления истощения ее. Смену признаков гипертиреоидного состояния на признаки гипофункционального выявляли у собак через 2—3 года после общего облучения или облучения верхней части туловища (накопление железой радиойода, содержание связанного с йодом белка в крови, реакция на тиреотропную стимуляцию). У части животных развилась микседема.
Аналогичная последовательность изменений функции коры надпочечников может быть прослежена после воздействия радиации. При лучевой болезни у собак вслед за усилением их функции первые признаки недостаточности коры надпочечников стали появляться спустя 3 мес. Наблюдались все возрастающие патологические изменения в надпочечниках по мере отдаления срока после лучевого воздействия. Э. Бетц первый обратил внимание на то, что состояние некоторых инкреторных функций после воздействия радиации такое, какое может наблюдаться при постоянном поддержании реакции напряжения, и при этом особое внимание было обращено на длительно сохраняющееся состояние гиперкортицизма.
Об отдаленных изменениях в эндокринных функциях у людей, переживших ядерный взрыв в Хиросиме и Нагасаки, много сообщалось. При этом подчеркивалась определенная однотипность этих изменений в различных эндокринных органах, нередко доходивших до стадии истощения. Известно, что значение дисгормональных расстройств особенно возрастает в период отдаленных лучевых последствий. Возникновение ряда опухолей эндокринных органов (в первую очередь щитовидной железы) в отдаленный период отнесено к наиболее радиочувствительному виду радиационных неоплазм. Характерно, что для развития этих опухолей необходим определенный оптимум дозы, который бы не приводил к гибели и позволил в течение длительного времени сохранять напряжение функций эндокринных желез.
Общую последовательность развития отдаленных изменений эндокринного статуса облученного организма можно представить так. У благополучно перенесших лучевое воздействие наступает полное клиническое выздоровление, несмотря на то, что в отдельных системах имеются остаточные повреждения и нарушения. Они не проявляются и могут никогда не проявиться, пока компенсаторно-приспособительные реакции организма находятся на адекватном уровне для поддержания состояния стабилизация здоровья. Однако в ряде случаев через определенный латентный период повышается функциональное состояние периферических эндокринных желез (щитовидная железа и, возможно, другие), в организм поступает все большее количество гормонов этих желез без явлений гормональных токсикозов. Организм не испытывает их избытка. Наоборот, имеются основания предполагать, что в этих условиях ткани-потребители гормонов испытывают их недостаток, так как дополнительное введение их в организм тоже не вызывало токсикоза и оказывалось во многих отношениях благоприятным. Длительное состояние гиперфункции желез компенсаторного плана в связи с дефицитом гормонов в конце концов может переходить в гипофункциональное состояние и истощение. Возрастающий со временем недостаток гормонов, т. е. срыв компенсации, создает благоприятные условия для клинического проявления остаточных и возникновения отдаленных радиационных последствии. Как показали наши исследования, такие последствия, как напряжение эритропоэза и сниженная иммунобиологическая реактивность, проявлялись в большей степени в те периоды, когда дефицит в гормоне щитовидной железы предполагался более выраженным.
В качестве примера возможного экспериментального подтверждения указанных положении сошлемся на опыт по изучению эритропоэза. Были получены следующие результаты: у крыс напряженность гемопоэза была более выражена через 4 мес., а не через 2 мес. после облучения. Свидетельством того, что через 4 мес. организм испытывал более выраженный недостаток в гормонах щитовидной железы, может, по-видимому, служить более высокая эффективность 30-суточного введения тиреоидина в малых физиологических дозах через 4 мес. после облучения, чем через 2 мес.
Наблюдения над людьми, перенесшими радиационное воздействие, показывают, что нередко следует говорить лишь о практическом выздоровлении. Различные факторы нерадиационной природы часто служат внешним поводом для проявления отдаленных радиационных последствий. Так, у лиц, подвергавшихся радиотерапии по поводу злокачественных заболеваний, отмечали, что толчком к появлению астенизации иногда служили внешние факторы нерадиационной природы. Они выводили больных из состояния компенсации.
Есть данные о том, что через 9 лет у большинства лиц, перенесших воздействие радиации ядерного взрыва, несмотря на восстановление здоровья, реакция их на всякое напряжение изменяется в сторону возможности появления хронических расстройств. Это связано с тем, что патогенетически более поздние проявления радиационного поражения являются в определенной мере универсальными и их выраженность зависит во многих отношениях от одних и тех же компенсаторно-приспособительных реакций, а срыву компенсации могут способствовать разные факторы.
Взаимно отягощающее влияние разных факторов на состояние щитовидной железы показано и на примере воздействия радиации и инфекции. Значительное повышение функции железы у крыс вызывали раздельно радиацией (через 4 мес. после облучения в дозах 7,5 и 8,5 Гр) и заражением газовой гангреной (исследовали морфологию железы, массу секретирующей тиреоидной ткани, содержание меченого тиреоглобулина в железе, массу железы). У крыс, которых заражали газовой гангреной через 4 мес. после воздействия радиации, гистологическое состояние щитовидной железы указывало на значительное усиление ее функции. Однако оно не сопровождалось увеличением массы железы, а содержание тиреоглобулина в железе было существенно ниже, чем у здоровых животных. Таким образом, инфекция, сама по себе вызывающая у здоровых животных гиперфункцию железы, привела у облученных крыс к состоянию, близкому к ее истощению.
При дефиците гормонов периферических желез (на примере недостаточности щитовидной железы) выявляется временная связь с рядом проявлений как радиационных последствий, так и нарушений, не имеющих отношения к радиации. Высказано мнение, например, о том, что ускоренное разрушение эритроцитов может быть не связано с качеством их, а обусловлено недостатком гормонов в организме. Известно, что при гипотиреозных состояниях продолжительность жизни эритроцитов уменьшается почти в 2 раза по сравнению с продолжительностью их жизни у здорового человека. Клиницисты-гематологи выделяют специальную форму анемии — тиреоривную анемию, развивающуюся на почве гипотиреоза и микседемы, основным лечением которой является введение в организм тиреоидных гормонов.
При этом для нас важным представляется тот факт, что эритроциты больных микседемой, перелитые здоровым реципиентам, стали иметь нормальную продолжительность жизни. Обнаружено также, что и при ряде других болезней, сопровождаемых укорочением продолжительности жизни эритроцитов, переливание их в здоровый организм обусловливало нормальную продолжительность их жизни (серповидноклеточная анемия, нефриты) и, наоборот, переливание эритроцитов от здоровых людей в организм больных укорачивало продолжительность их пребывания в русле крови (ревматоидный артрит, острые инфекционные процессы).
Все это свидетельствовало о том, что основной и непосредственной причиной укорочения продолжительности жизни эритроцитов в данных случаях было не ухудшение их качества, а не связанные с ними факторы организма. Эти факторы могут быть разными, но в абсолютном большинстве случаев они непосредственно связаны с селезенкой, с гиперфункциоиальным ее состоянием. Для нас важно, что действие этих факторов по времени совпадает с недостаточностью периферических эндокринных желез.
Указанные отдаленные радиационные последствия, в том числе и злокачественные, не отличаются от таких же заболеваний, вызванных другими факторами.
Более того, они характерны для возрастной патологии и часто возникают только тогда, когда возраст организма приблизился к периодам проявления аналогичной спонтанной патологии (дегенеративно-дистрофические нарушения, склеротические, циррозы печени, нефросклерозы, неопластические заболевания и т. п.). В этот период также наблюдаются определенные возрастные изменения в эндокринной системе. Изложенное породило специальные направления исследований, таких, например, как «Старение, климакс и рак» [Дильман, 1968] и «Эндокринологическая онкология» [Дильман, 1974].
Возрастные изменения и гормональная регуляция
Два обстоятельства в исследовании возрастных изменений обращают на себя внимание: 1) некоторая общность проявлений отдаленной радиационной и возрастной патологии; 2) взаимосвязь многих проявлений возрастной патологии с состоянием эндокринных желез.
Старение — процесс универсальный, характеризующийся своими внутренними закономерностями и захватывающий все системы организма. Появляется функциональная неполноценность клеток разного типа, уменьшается их число и на оставшиеся ложится повышенная функциональная нагрузка. Существенно изменяются реактивность клеток, их чувствительность к регуляторным влияниям. Возрастные изменения затрагивают процессы генерации энергии и ограничивают ее возможности. На примере мышечных волокон показано, что при старении наступает перераспределение различных путей энергетического обеспечения клеток, все большее значение начинают приобретать гликолитические процессы. Уменьшается число белковых рецепторов на мембране, и изменяется их состояние, зависимое от фосфолипидного окружения.
В.В. Фролькис считает, что частичная дерецепция клеток при старении нарушает межклеточные взаимоотношения и участие клеток в системных, общерегуляторных реакциях, что является важным механизмом расстройства их функции. Многие изменения в клетках в процессе старения сходны со сдвигами в денервированной ткани. Ослабляется нервный и гуморальный контроль деятельности клеток.
Однако в ответ на возрастание изменения и падение способности клеток отвечать адекватной реакцией развиваются в первый период старения достаточно эффективные компенсаторно-приспособительные и адаптивные реакции. Так, известно, что в этот период происходит возрастание чувствительности клеток некоторых органов к катехоламинам, может наблюдаться даже рост содержания в них цАМФ. Тем не менее, общие адаптационные возможности организма и его важнейших систем к старости значительно сужаются. Возрастные изменения в таких системах, как сердечнососудистая, дыхательная и пищеварительная, способствуют развитию явлений кислородного голодания организма и недостатка ряда субстратов обмена. Наиболее ярко видны возрастные изменения в иммунной и эндокринной системах.
Прогрессивное уменьшение массы лимфоидной ткани после рождения и на протяжении всей последующей жизни хорошо известно. Это заметно по уменьшению тимуса, лимфатических узлов, селезенки и числа лимфоцитов в крови. Наблюдения на людях и в экспериментах на животных дали много доказательств связи между болезнями старения и нарушением иммунных функций. В эту группу заболеваний включают воспалительные заболевания, васкулярные и особенно кардиоваскулярные и почечные заболевания, так называемые дегенеративно-сосудистые заболевания, аутоиммунные заболевания, предрасположенность к инфекционным и злокачественным заболеваниям и амилоидоз.
| Возраст | Число лиц с аутоантителами,% | Средний титр |
| 12-14 лет 17-22 года 46-59 лет 60-75 лет | Нет 13 38 50 | Нет 1:2 1:6 1:10 |
С возрастом лимфоидная ткань частично заменяется грубоволокнистой соединительной тканыо, количество лимфоцитов уменьшается, их пролиферация ослабляется. Тем не менее, увеличивается количество антител к собственным тканям организма. Это четко было показано на примере роста с возрастом мужчин числа комилементсвязывающпх аутоантител к ткани яичка (В.М. Шубник).
Старение характеризуется иммунологической недостаточностью, которая является следствием развития дефектов всех этапов иммуногенеза. Происходит дефицит и клеточного (Т-система лимфоцитов), и гуморального (В-система лимфоцитов) иммунитета. С возрастом постепенно уменьшается пул стволовых клеток в кроветворных тканях, снижается миграция стволовых элементов из костного мозга. Взаимодействие Т- н В-лимфоцитов ухудшается, их пролиферация угнетается (Р.В. Петров).
Однако с возрастом и под влиянием разнообразных внешних воздействий усиливаются аутоиммунные процессы. Усиление аутоиммунных процессов при старении обычно находится в обратной зависимости от способности к нормальному иммунному реагированию. Подчеркивается, что при старении частота возникновения рака значительно возрастает, что совпадает по времени с развитием возрастной иммунологической недостаточности.
Необходимо также подчеркнуть некоторые общие особенности иммунной системы:
1) иммунологическая система является очень чувствительной к возрастным изменениям и внешним воздействиям;
2) восстановление нарушений в системе иммунитета происходит с большими затруднениями, неполноценно и в очень отдаленные сроки;
3) основным источником, определяющим клеточное содержание системы иммунитета, является поступление коммутированных в сторону лимфопоэза кроветворных клеток;
4) высокая чувствительность системы иммунитета к возрастным изменениям и повреждающим воздействиям, затруднения в ее восстановлении могут быть связаны с недостаточным пополнением пула предшественников лимфопоэза, что обусловлено приматом дыхательной функции крови над функцией иммунологической защиты.
Материал показывает также, что в первичных процессах канцерогенеза система иммунитета непосредственного участия, по-видимому, не принимает. Ее функции в этом плане сводятся к уничтожению малигнизированных клеток и недопущению их прогрессии. В случае ослабления функции иммунологического надзора приобретается предрасположенность к опухолевой прогрессии.
Однако возрастное увеличение аутоагрессии против собственных клеток и тканей, как будет показано далее, может быть элементом цепи событий, создающих условия, также способствующие тому, чтобы клетки изменялись в сторону приобретения черт злокачественности.
На связь многих проявлений отдаленной радиационной патологии с состоянием эндокринной системы мы уже также обращали внимание. Воздействие радиации ускоряет и увеличивает вероятность развития возрастной патологии, включая злокачественные заболевания. Связь многих проявлений возрастной патологии с эндокринологическими изменениями наиболее обстоятельно изучена В.М. Дильмапом. Он обратил внимание на закономерное повышение порога чувствительности гипоталамуса к периферическим гормонам и глюкозе на протяжении всей жизни человека. Отмечено изменение с возрастом числа рецепторов инсулина на плазматической мембране клеток гипоталамуса. В.М. Дильмапом считает, что сохранение эффективно работающего механизма обратной связи возможно только в том случае, если параллельно происходит и компенсаторное усиление периферического звена системы гомеостата. Соответственно в энергетической системе гомеостата (регуляция энергетического обмена организма) компенсация осуществляется за счет повышения продукции инсулина в поджелудочной железе, в репродуктивной системе гомеостата — за счет увеличения суммы классических и неклассических стероидов в яичниках, в адаптационной системе гомеостата — за счет увеличения продукции кортизона корон надпочечников. Гормоны периферических эндокринных желез участвуют в торможении соответствующих центров гипоталамо-гипофизарпон системы и уменьшают тем самым выделение тройных гормонов гипофиза, регулирующих активность периферических желез.
Вследствие развития с возрастом этих реакций компенсации происходят нарушения гормонально-метаболического равновесия. Среди них: ослабление торможения глюкозой секреции гормона роста, понижение чувствительности к действию инсулина, компенсаторная гиперин-сулипемия, увеличение массы тела, снижение толерантности к глюкозе, повышение в крови уровня жирных кислот, холестерина, триглицеридов, относительное увеличение продукции оксистероидов, повышение секреции тотальных гонадотропинов и тотальных фенолстероидов и др. Так, в течение каждого десятилетия жизни у женщин наблюдается примерно двукратное увеличение выделения гонадотропинов по сравнению с предыдущим десятилетием. Многие из этих признаков нарушения гомеостата организма обнаруживаются и у больных с различными видами злокачественных заболеваний.
Предполагают, что такой гормонально-метаболический фон формирует условия, обеспечивающие развитие возрастной патологии, и способствует возрастному нарастанию частоты многих видов рака. В особую группу патологии выделяют болезни компенсации, связанные с повышением гипоталамического порога, целый ряд заболеваний и состояний: возрастную гиперхолестеремию и атеросклероз, ожирение, гипертоническую болезнь, некоторые формы сахарного диабета, болезнь Иценко—Кушинга, базедову болезнь, аменорею, полигормональные кровотечения, гипофизарный экзофтальм, полицитемию и некоторые формы злокачественных заболеваний. Эта обширная группа патологических процессов характеризуется отчетливой возрастной динамикой, частым сочетанием ряда признаков у одних и тех же лиц, лечебным действием средств, ингибирующих гипоталамо-гипофизарную систему.
В.М. Дильман отмечает, что эндогенный механизм возрастного повышения порога чувствительности гипоталамуса формирует с той или иной скоростью болезни компенсации независимо от степени влияния факторов внешней среды. В то же время он указывает на то, что многие внешние воздействия, способствующие развитию возрастной патологии (стресс, избыточность питания, частично ионизирующая радиация и канцерогены), оказывают свое патогенное воздействие путем интенсификации механизма повышения гипоталамического порога (сам механизм остается нераскрытым). Поэтому в реальных условиях создается интегральное влияние внутренних и внешних факторов формирования болезней компенсации и на их основе ряда злокачественных заболеваний. Синдром понижения сопротивляемости к инфекции возникает как в процессе нормального старения, так и под влиянием тех внешних воздействий, защита от которых осуществляется через напряжение адаптационного гомеостаза.
В.М. Дильман рассматривает варианты гомеостатической недостаточности, в основе которых — повышение порога чувствительности гипоталамуса. Для нас важно, что изменение эффективности и искажение регуляторных связей между разными органами создают предпосылки для развития ряда патологических состояний. Часто в их основе лежат реакции компенсации и сверхкомпенсации в ответ на ослабление или искажение межсистемных взаимодействий. Имеются яркие примеры развития не только возрастных, но и опухолевых процессов.
Так, Ю.И. Москалев и В.Н. Стрельцова, анализируя возможные механизмы развития злокачественных опухолей щитовидной железы после воздействия ионизирующих излучений, рассматривают следующую последовательность событий: радиационное воздействие -»- радиационное повреждение щитовидной железы->-снижение секреции тиреоидных гормонов->-понижение уровня тиреоидных гормонов в крови - гиперфункция гипофиза и избыток выработки тиреотропного гормона-гиперплазии сохранившейся ткани щитовидной железы->-гормонозависимая аденома щитовидной железы-«-гормононезависимая (злокачественная) опухоль щитовидной железы.
В указанной последовательности событий опухоль вызывается не воздействием радиации и не радиационно-генетическим повреждением тканей железы, а непосредственной избыточной стимуляцией ее тиреотропным гормоном гипофиза. Возникшая гиперплазия оставшейся функционально способной ткани железы и стала главным фактором, приведшим к опухолевому процессу.
Такую последовательность событий подтверждают и массовые опыты на чистопородных собаках, которых облучали в очень малых дозах (0,16 пли 0,83 Гр) перед или в ближайшие сроки после рождения. Ряд авторов проследили все этапы развития, начиная с атрофии железы с последующими очагами гиперплазии фолликулярных клеток до неоплазии и образования эктопических опухолей тиреоидной ткани в отдаленные сроки после облучения. Вот последовательности событий, отмеченные этими авторами:
1) введение в организм больших доз радиоактивного йода->-концентрация йода в щитовидной железе-»-ингибирование функций щитовидной железы - напряжение тиреотропной функции гипофиза->-опухоль гипофиза;
2) локальное облучение яичников->-фолликулярные кисты яичников->-дисгормональные опухоли матки, молочной железы.
Заключение
Во всех указанных случаях неопластические процессы возникали из клеток, непосредственно не подвергавшихся, воздействию радиации.
Материалы позволили предполагать повышение пострадиационного порога чувствительности органов-мишеней для гормонов периферических желез, что снижает эффективность действия этих гормонов и влечет за собой цепь последующих изменений. Среди них — компенсаторное повышение функциональной активности этих желез, чреватое переходом в патологическую гиперактивность или истощение. Следовательно, и в этих случаях можно говорить о возможном развитии болезней компенсации и об увеличении частоты возрастной патологии. При этом главное внимание мы обращали на исследование скрытой патологии со стороны системы крови.
Таким образом, изменение порога чувствительности к соединениям, осуществляющим прямую и обратную связь, с возрастом или под влиянием внешних воздействий может возникать не только в гипоталамусе, но и в периферических тканях органов-мишеней для действия гормона. По-видимому, они могут возникать в любом звене общей гомеостатической системы: гипоталамусе, гипофизе, периферических эндокринных железах и в тканях органов-мишеней. Так, снижение эффективности действия гормона вследствие меньшей чувствительности воспринимающих тканей показано как следствие воздействия радиации на примере чувствительности репродуктивной системы к гонадотропному гормоиу. Реакция щитовидной железы на введение тиреотропного гормона гипофиза с возрастом существенно понижается.
В таком случае снимается уникальность гипоталамуса как единственного звена гомеостатической системы, в котором происходит повышение (в более общем виде — изменение) порога чувствительности. Однако гипоталамус занимает верховное иерархическое положение в системе гомеостата, и потому изменения в нем будут иметь наибольшие последствия.
Молекулярные механизмы неотвратимого возрастного или ускоренного под влиянием внешних воздействий изменения порога чувствительности элементов системы гомеостата организма совершенно неизвестны. Совокупность имеющихся данных позволяет предполагать их связь с различными особенностями нарушений клеточного цикла пролиферирующих тканей и ошибок синтеза белка в активно функционирующих тканях.
| Литература - Другое (клиника, диагностика, лечение некоторых форм) | |
|
Этот файл взят из коллекции Medinfo http://www.doktor.ru/medinfo http://medinfo.home.ml.org E-mail: medinfo@mail.admiral.ru or medreferats@usa.net or ... Миграцию и хемотаксис фагоцитирующих клеток можно оценить при использовании агарозных тестов; поглотительную способность лейкоцитов измеряют по захвату ими различных частиц ... Блок 1 - Отсутствуют стволовые клетки, поэтому организм практически беззащитен. |
Раздел: Рефераты по медицине Тип: реферат |
| Мононуклеарный онкогенез | |
|
МОНОНУКЛЕАРНЫЙ ОНКОГЕНЕЗ Введение злокачественный мононуклеарный заболевание лечение Онкогенез (греч. onkos - масса, нарост, опухоль; genesis ... Из всех клеток системы крови на роль "общего начала" или клетки-предшественницы первичной стволовой злокачественной клетки солидных опухолей могут претендовать только Мононуклеары ... Мононуклеарная фракция в системе крови - это клетки, выделенные из костного мозга или периферической крови посредством отделения от эритроцитов, тромбоцитов и гранулоцитов на ... |
Раздел: Рефераты по медицине Тип: книга |
| Радиационная безопасность при эксплуатации и ремонте оборудования ... | |
|
Федеральное агентство по атомной энергии Федеральное государственное унитарное предприятие "Российский государственный концерн по производству ... осуществлять контроль за дозами облучения, полученными подчиненным персоналом, регулярно информировать его о состоянии радиационной обстановки на рабочих местах и индивидуальных ... При этом в крови резко уменьшается количество белых кровяных телец - лейкоцитов (в значительной степени уменьшаются защитные силы организма в борьбе с инфекцией), кровяных ... |
Раздел: Рефераты по безопасности жизнедеятельности Тип: учебное пособие |
| ... при уролитиазе кошек и собак в условиях мегаполиса и контроль за их ... | |
|
Министерство сельского хозяйства Московская Государственная академия ветеринарной медицины и биотехнологии Дипломная работа "Особенности ... Недостаточность функции паращитовидных желез сопровождается понижением количества кальция в крови, и наоборот, избыточный выброс гормона вызывает повышение концентрации кальция в ... 2) Лейкоциты крупнее эритроцитов (10-12 микрон), но значительно меньше эпителиальных клеток. |
Раздел: Рефераты по ветеринарии Тип: реферат |
| Общая нозология. Типовые патологические процессы | |
|
КОНТРОЛЬНАЯ РАБОТА ПО ОБЩЕЙ ПАТОЛОГИИ РАЗДЕЛ 1. ОБЩАЯ НОЗОЛОГИЯ Тема 1. Патогенез (морфогенез и функциогенез) болезни Этиотропная (каузальная) терапия ... Рак возникает под влиянием химических веществ, ультрафиолетового облучения, гормонов, вирусов, радиации. В норме кровь имеет характер суспензии клеток крови в жидкой части ее, определяется величиной отрицательного заряда эритроцитов и тромбоцитов, соотношением белковых фракций плазмы ... |
Раздел: Рефераты по медицине Тип: контрольная работа |