Реферат: Основные понятия молекулярной биологии
Реферат на тему:
Основные понятия молекулярной биологии
2009
Особенности атомов четырех элементов таблицы Менделеева, составляющих основу всех биологических молекул: водорода, углерода, азота и кислорода.
Водород. Номер 1 в таблице Менделеева. Малое по размеру ядро несет единичный положительный заряд. Электрон находится на первой, близкой к ядру орбите. Атом водорода легко теряет электрон, образуя положительный ион Н+. Говорят, что атом водорода является хорошим донором электрона, переходящего к другим атомам. Ион Н+ играет важную роль в биохимических реакциях.
Углерод (№ 6). Заметно большее, чем у водорода ядро имеет заряд +6. Соответственно, электронов тоже 6. Два электрона, первой, ближайшей к ядру оболочки не способны участвовать в образовании химических связей. На второй, «валентной» оболочке — 4 электрона. Они могут находиться в двух состояниях. Первое — когда два электрона из четырех спарены (определенным образом связаны между собой) и потому не могут принимать участия в образовании химических («ковалентных») связей с другими атомами. В этом состоянии углерода двухвалентен, как например, в молекуле СО. В образовании связей участвуют только два не спаренных электрона второй оболочки. Ковалентную связь между двумя атомами образуют два валентных электрона — по одному от каждого из атомов. Эти электроны спариваются и обращаются по общей орбите вокруг обоих ядер. В этом «совладении» электронами и состоит химическая связь. Если электроны спарены на орбите одного из атомов, то они в химической связи участвовать не могут.
Однако, как раз у углерода спаренные электроны наружной оболочки легко разъединяются и приобретают способность участвовать в образовании ковалентных связей. В этом состоянии углерод четырехвалентен (например, COg). Четырехвалентный углерод, связываясь с четырьмя другими атомами (например, в молекуле метана СН4), образует очень устойчивую пространственную конфигурацию тетраэдра, в центре которого находится ядро углерода.
Добавим, что ядро углерода прочно удерживает все четыре валентных электрона внешней оболочки. Поэтому углерод не склонен образовывать ион.
Азот (№ 7). В нормальном состоянии имеет на внешней оболочке пять электронов, из которых два спарены. Поэтому в соединениях азот чаще- всего трехвалентен (например, в аммиаке — NH3). Однако образование тетраэдрической структуры настолько выгодно, что атом азота охотно отдает один из спаренных электронов и превращается в четырехвалентный положительно заряженный ион, который соединяется ковалентно с четырьмя другими атомами, например в ионе аммония NH3.
К взаимоотношениям нормального, трехвалентного азота и водорода стоит присмотреться внимательнее.
Во-первых, партнерство маленького водорода с более крупным азотом далеко не равноправное. Общая пара электронов должна обегать оба ядра. Но в окрестностях азота она проводит куда больше времени, чем около ядра водорода. Это означает, что атом азота получает небольшой дополнительный отрицательный заряд (его обозначают 5-), а водород оказывается в такой же мере положительно заряженным (б+). Происходит смещение заряда . Значки 5 на этом рисунке не имеют количественного смысла, а лишь обозначают малую величину, заведомо меньшую 1. Две другие химические связи азота не сильно влияют на это смещение.
Во-вторых, из-за наличия на внешней орбите атома азота спаренных электронов этот атом за счет их отрицательного заряда дополнительной, хотя и очень небольшой, силой удерживает около себя атом водорода.
Эти особенности взаимоотношений азота и водорода играют в живой природе очень важную роль.
Кислород (№ 8). В нормальном состоянии на внешней орбите атома находится шесть электронов, из которых четыре спарены, и только два могут участвовать в образовании химических связей. Кислород двухвалентен. Помимо этого, он обладает уникальной особенностью: весьма склонен отнимать у других атомов электрон и включать его в состав своей внешней оболочки, превращаясь в отрицательный ион. В этом смысл процесса окисления. Захваченный электрон спаривается с одним из двух прежде свободных электронов кислорода и его ион оказывается одновалентным. Говорят, что кислород обладает сродством к электрону. Это обусловлено тем, что присоединивший седьмой электрон к своей внешней электронной оболочке ион кислорода приближается к очень устойчивой конфигурации инертного атома неона. Азот и кислород в этом плане являются антиподами. Азот может служить донором электрона, а кислород является его акцептором»
Сродство кислорода к электрону проявляется и тогда, когда оба его свободных электрона заняты в образовании ковалентных связей. Кислород «стремится» удержать около себя спаренные электроны этих связей, оттягивая на себя таким образом электронную плотность и получая некоторый отрицательный заряд.
Кроме того кислород, имея на внешней орбите спаренные электроны (даже две пары), способен, подобно азоту притягивать оказавшийся поблизости дефицитный по электронной плотности атом водорода.
Описанные особенности атомов азота и кислорода позволяют понять природу слабых сил взаимодействия, играющих ключевую роль как в образовании пространственной структуры белков и ферментативном катализе, так и в спаривании нитей ДНК. Рассмотрим три главных типа таких сил.
1. Ионная связь. Такая связь может возникнуть при сближении отрицательного иона кислорода (например после диссоциации водорода от карбоксильной группы) и положительного иона четырехвалентного азота, как показано на рис. 3. Не интересующие нас пока остальные участки молекул изображены в виде прямоугольников).
Расстояние между ядрами ионов кислорода и азота около ЗА, а сила взаимного притяжения такова, что для отрыва их друг от друга требуется затратить 0,95 килокалорий на каждый моль ионных пар. Для разрыва же ковалентной химической связи (например, между двумя атомами углерода) требуется почти в сто раз больше энергии — около 85 ккал/моль.
2. Водородная связь. Образование так называемой «водородной» связи обусловлено указанными выше явлениями:
поляризацией связи N—Н
притяжением обедненного электронами водорода к атому азота или кислорода, имеющим на внешней орбите спаренные электроны.
Расстояния между ядрами двух азотов или азота и кислорода такие же, как при ионной связи — около ЗА. Ядро водорода как бы зажато между почти соприкасающимися внешними электронными оболочками N и N или N и О. Для разрыва водородной связи требуется около 0,7 ккал/моль — это тоже слабая связь.
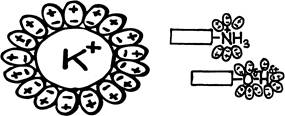
3. Гидрофобная связь. Это, по существу говоря, не связь, а «невольное» объединение молекул, отторгнутых молекулами воды. Гидрофобные молекулы «не любят» воду — растворяются в ней очень плохо. Растворимость в воде означает склонность вещества распадаться на отдельные молекулы, каждую из которых окружает гидратная оболочка — слой «прилипших» молекул воды. «Прилипание» воды (смачивание) происходит благодаря притяжению поляризованных молекул воды (диполей) к ионам или заряженным участкам растворяемых молекул (рис1). Поляризованность молекулы воды, если учесть, что в реальности оба водорода действительно находятся по одну сторону от атома кислорода.
|
|
|
|
| Рис.1 |
| Рис.2 |
Если крупная молекула имеет на поверхности заряженную или поляризованную группу атомов, то вокруг нее тоже будут образовываться «местные» гидратные оболочки (рис. 2). Этого может быть достаточно для придания всей молекуле определенной (худшей, чем для чистого иона) растворимости в воде. К первому слою «прилипших» молекул воды такими же силами присоединяются еще слои молекул воды, образуется своего рода «шуба», которая «втягивает» молекулу в водное окружение, даже если остальные участки молекулы с водой не взаимодействуют.
Естественно, что сами диполи молекул воды электростатически и водородными связями притягиваются друг к другу. Это обусловливает, в частности, некоторую трудность испарения воды (по сравнению, например, с эфиром и спиртом). Вода «стремится» упорядочить расположение своих молекул по объему так, чтобы между ними образовалось максимальное число связей. Чем больше таких связей, тем меньше потенциальная энергия всей системы (ведь для разрыва связей, например испарения воды, надо подводить энергию нагревания). Любая подвижная система стремится занять положение, отвечающее минимуму потенциальной энергии. Тепловое движение молекул воды препятствует этому. Максимальное упорядочивание достигается лишь при 0°С, когда вода кристаллизуется в лед.
Гидрофобные молекулы образуются незаряженными и неполяризованными атомами и группами атомов. Они не связывают воду, не смачиваются. Таковы, например, алифатические цепочки вида СНз—CH2—CH2—... да и сами образующие их группы CH3— и —CH2— . Высока гидрофобность ароматических молекул: бензола (C6H6), фенола, толуола и других, в состав которых входят разного рода замкнутые циклы с сопряженными связями. Вода такие молекулы как бы «не замечает». Но, оказавшись в воде, несмачиваемые молекулы занимают место и мешают молекулам воды образовать максимально возможное при данной температуре число связей. Помехи уменьшатся, если гидрофобные молекулы соберутся в тесные кучки (капли), как показано на рис. 3, где кружками изображены гидрофобные молекулы, а зачерненными — молекулы воды.

Рис.3
Именно так вода «выталкивает» гидрофобные молекулы, точнее — сохраняет их случайное объединение. И хотя сами гидрофобные молекулы никак не взаимодействуют друг с другом, конечный результат выглядит так, будто они притягиваются некими силами, которые и условились называть «гидрофобными связями». Чтобы эти молекулы разъединить (растворить), надо затратить энергию на разрыв связей между молекулами воды, расчистить места для распределения гидрофобных молекул по всему объему. (Именно поэтому жир относительно легко смывается горячей водой).
Энергия разрыва таких связей, считая на моль гидрофобного вещества (при условии избытка воды) заметно больше, чем в случае ионной или водородной связи, а именно — около 2,4 ккал/моль. Но и она мала по сравнению с ковалентной связью. Так что гидрофобная связь тоже слабая.
Белковая глобула. Аминокислоты
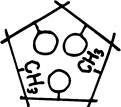
Мы уже знаем, что все белки состоят из комбинации 20-ти аминокислот. Их можно получить из любой смеси белков гидролизом в 6М соляной кислоте при 110°Св течение суток без доступа воздуха (в запаянной ампуле). Методом хроматографии, — с которым мы в свое время познакомимся, — все 20 аминокислот, находящиеся в гидролизате можно разделить между собой. Упоминалось, что аминокислоты, образуя белок, выстраиваются в прочные цепи и что они обладают разного рода свободными, химическими группами атомов. Первое обстоятельство обеспечивается наличием во всех аминокислотах трехзвенной цепочки атомов, показанной на рис. 8. Прямоугольник и ромб там обозначают некие боковые ветви, строением которых аминокислоты отличаются друг от друга.
W-CH-COOH
Очевидно, в боковых ветвях и следует искать свободные группы атомов. Этим мы займемся немного позже. А сейчас посмотрим на доступные нашему вниманию трехчлены. На одном конце у них стоит аминогруппа (NHg), на другом — карбоксил (СООН), характерный для карбоновых кислот. Отсюда и название — аминокислоты.
Теперь приступим к рассмотрению боковых ветвей аминокислот. Для упрощения рисунка теперь уже цепочку трехчлена будем изображать в виде прямоугольника. Под ним — полное и сокращенное наименование аминокислоты, а во второй строчке — условное однобуквенное ее обозначение и молекулярный вес (дальтон). По химическим особенностям боковых ветвей аминокислоты можно разбить на несколько групп.
1. Аминокислоты, несущие электрический заряд (в воде):
а) основные: заряд положительный
б) кислые: заряд отрицателен
У аспарагина и глутамина в образовании водородных связей участвуют аминогруппы и атомы кислорода, а у серина и треони-на в этой роли выступают ОН-группы.
3. Аминокислоты, склонные к образованию гидрофобных связей:
а) алифатические;
б) ароматические
Заметное ультрафиолетовое поглощение растворов белков при 280 нм обусловлено главным образом поглощением трипто-фана, в меньшей степени — тирозина и совсем незначительно — фенилаланина.
4. Серусодержащие аминокислоты Эти аминокислоты не участвуют в образовании слабых связей. Для окончательной фиксации белковой глобулы неплохо было бы ее закрепить более прочными химическими «сшивками». Эту задачу берут на себя пары цистеинов. Сблизившись группами SH и вступив в реакцию окисления они образуют ковалентную связь между атомами серы Такую связь называют дисульфидным мостиком. Во многих белках дисульфидные мостики отсутствуют, в других их немного (3-6), но бывают и исключения. Например, в сывороточном альбумине быка их 17. Дисульфидную связь можно разорвать добавлением восстанавливающего агента, например, бета-мер-каптоэтанола (HS—С2Н5ОН).
Метионин, как мы увидим, играет особую роль в инициации синтеза белка.
5. Последняя аминокислота Пролин (Про) имеет особую структуру. Нарисуем ее формулу в полном виде . Собственно говоря, это не аминокислота, а иминокислота,так как вместо аминогруппы (NH3) в трехзвенной цепочке, стоит иминог-руппа (NH). Но это не мешает образовывать пептидную связь, имеющую вид, изображенный на рис. 17. Трехзвенная цепочка пролина не прямолинейна, а образует некий угол. Это обеспечивает изгиб цепи белка в том месте, где в ней находится пролин.
Все белки имеют одинаковый элементарный химический состав. Между тем, набор аминокислот у различных индивидуальных белков разный. Противоречие снимается, если вспомнить, что белковые цепи содержат сотни аминокислот. Суммарный состав усредняется до примерно следующих величин: 52% С; 22,5% О; 16% N; 7% Н; 1,5% S, по 1% Р и Fe (по весу). Фосфор и железо входят в состав не всех белков.
Образование белковой глобулы.
Для упрощения рисунков введем условные обозначения групп аминокислот.
Процесс сворачивания белковой цепи после ее синтеза происходит в два этапа.
I этап. Исходно удаленные друг от друга гидрофобные аминокислоты постепенно сближаются и образуют сильно гидрофобные зоны .
Такие зоны, по возможности, стремятся еще сблизиться и образовать гидрофобное ядро глобулы. Ввиду ограниченности гибкости цепи не все гидрофобные аминокислоты попадут внутрь. Некоторые гидрофобные боковые ветви окажутся на поверхности глобулы. Однако преобладать там (кроме особых случаев, например белков мембран) будут боковые ветви гидрофильных аминокислот. Это обеспечит хорошую смачиваемость, а значит — растворимость белка в водной среде.
II этап. Хотя глобула прошла этап первоначального формирования и вчерне свернулась, она еще рыхлая. Толчки молекул воды могут вызвать взаимные перемещения участков глобулы. В ходе этих перемещений сближаются активные группы боковых ветвей аминокислот, способные образовать дополнительные слабые (ионные и водородные) связи. Они фиксируют структуру глобулы. Если фиксации не происходит, то первоначально образовавшиеся гидрофобные зоны и все внутреннее ядро могут претерпеть одну или несколько реорганизаций, пока не будет найдена оптимальная структура, которую смогут зафиксировать ионные и водородные связи. Эти связи слабы, но их много, и глобула приобретает не только окончательную конфигурацию, но и достаточную прочность. Если же функция белка требует большей жесткости, то наследственно задается такое расположение цистеинов в первичной структуре белка, что в завершающей фазе формирования глобулы они оказываются друг против друга и прочно «сшивают» дисульфидными мостиками глобулу.
Как бы сложно не была свернута белковая глобула, она образована линейным полимером и потому имеет два конца — две концевых аминокислоты. Из способа соединения аминокислот, показанного на рис. 8, следует, что концевыми химическими группами всего белка будут: с одной стороны аминогруппа — NH3, с другой — карбоксил —СООН. Соответственно принято называть эти концы белковой молекулы: «N-конец» и «С-конец».
Расположенные на поверхности боковые ветви аминокислот определяют взаимоотношения белка с внешней средой. Электрически заряженные, поляризованные и склонные к образованию водородных связей активные группы гидратируются и тем самым, как уже упоминалось, обеспечивают растворимость белка в воде. Однако такие группы могут образовывать ионные связи и между разными глобулами, вызывая их агрегацию и вьшадание белка в осадок. Во избежание этого следует растворять белки не в чистой воде, а в слабом солевом растворе: ионы солей экранируют заряженные группы аминокислот (такая же солевая среда существует и внутри живых клеток). Иногда к раствору белка приходится добавлять (и в значительных количествах) мочевину, 0===C(NHg)g. Она способствует разрыву водородных связей между глобулами, замыкая эти связи на себя.
Необходимо разъяснить и природу явления «высаливания» — выпадания белков в осадок в крепких солевых растворах, например, сульфата аммония, (NH3SO4). Ионы солей активносвязывают воду, формируя собственные гидратные оболочки. Если таких ионов очень много, свободной воды не хватает для полной гидратации белков.
Второй важной характеристикой белков является электрический заряд. Суммарный заряд белковой глобулы равен алгебраической сумме положительных и отрицательных зарядов ионизированных боковых ветвей аминокислот, лежащих на поверхности белка. Ветви таких же аминокислот, оказавшихся внутри него, либо взаимно нейтрализованы в ионной связи, либо находятся в неионизованной форме из-за отсутствия воды внутри глобулы.
Боковые ветви аминокислот, расположенных на поверхности белковой глобулы, обеспечивают создание активного центра фермента. Рентгеноструктурньш анализ показал, что активный центр представляет собой впадину на поверхности белка, форма которой соответствует конфигурации молекулы (или молекул) субстратов ферментативной реакции. В строго определенных местах этой впадины располагаются активные группы аминокислот, ответственные за образование нескольких слабых связей с субстратом, удерживающих его во впадине и обеспечивающих правильную ориентацию реагирующих молекул .
Роль слабых связей внутри белковой глобулы показана в опытах по денатурации и ренатурации ферментов. В ряде случаев осторожным нагреванием, незначительными изменениями кислотности среды и другими воздействиями удается развернуть белковую глобулу (что подтверждают физические методы контроля). Фермент при этом «денатурирует» — утрачивает каталитическую способность. При постепенном восстановлении благоприятных условий глобула белка самопроизвольно и правильно сворачивается, ферментативная активность появляется вновь — происходит «ренатурация».
Литература
Авдонин П.В., Ткачук В.А, Рецепторы и внутриклеточный кальций.1994.-Наука, Москва. - С. 29-42.
Авцын А.П., Жаворонков А.А., Риш М.А., Строчкова Л.С. Микроэлементы человека, Медицина. М. -1991.
Анестиади В.Х., Нагорнев В.А. О пато- и морфогенезе атеросклероза. Кишинев, Медицина. -1985.-С.92.
Антонов В.Ф. Липиды и ионная проницаемость мембран. -М.: Медицина, 1982.
Аронов Д.М., Бубнова Н.Р., Перова Н.В. и др. Влияние ловастатина на динамику липидов и аполипротеидов сыворотки крови после максимальной физической нагрузки в период пищевой липемии у больных ИБС//Кардиология, -1995. -Т.35. -N 31. -С.38-39.
Атаджанов М.А., Баширова Н.С., Усманходжаева А.И. Спектр фосфолипидов в органах-мишенях при хроническом стрессе //Патологич. физиология и эксперим. терапия. -1995, -N 3: -С.46-48.