Реферат: Поведение животных при внутривидовых взаимоотношениях
Поведение животных при внутривидовых взаимоотношениях
Репродуктивный комплекс поведения включает в себя все то, что связано с размножением животных, и потому он имеет огромное значение для популяции вида, обеспечивает его существование во времени, связь поколений, микроэволюцию и соответственно гомеостаз популяции. В этот комплекс включено большое количество разных экологических актов и программ.
Перед тем, как приступить к изучению и описанию, нужно отметить, что в репродуктивном комплексе преобладают внутривидовые взаимоотношения. Наиболее эволюционно стабильные. Соответственно здесь преобладающими, но не единственными, есть видовые стереотипы, которые базируются, в основном, на врожденной основе.
В репродуктивном комплексе поведения каждого вида можно отметить несколько периодов, каждые из них имеет свою программу и следует один за один в виде цепочки, который подчиняется собственным сигналам как экзогенного, так и эндогенного характера.
Репродуктивная сигнализация
Здесь, скорее всего, все начинается из той сигнализации, с помощью которой особи разного пола, но одного вида, обнаруживают, находят и узнают о физиологической готовности к размножению особей другого пола. Эта сигнализация имеет очень большое значение для развития и существования вида, популяции. Она базируется на наследственно фиксированных особенностях видоспецифического поведения, которые изучает этология. Значение этой сигнализации весьма многогранно. Прежде всего, она дает возможность определять видовую принадлежность партнера, в значительной степени исключает межвидовую гибридизацию и сохраняет, таким образом, вид как морфофункциональную стабильность. Иными словами, если два сходных по своей морфологии вида будут иметь совпадающий ареал, то их скрещиванию и появлению бесплодных межвидовых гибридов обычно будут препятствовать «этологические преграды». Это своего рода коды, которые передаются типичными для вида звуками, запахами, окраской, телодвижениями и мимикой. При этом, хотя у животных, принадлежащих к самым разным классам можно видеть принципиально много сходного, но у каждого вида есть свой характерный набор видовых брачных сигналов, которые могут также несколько различаться и в различных участках видового ареала. Брачные сигналы дают возможность встреч и контактов, созревших и созревающих особей разного пола, выбора ими наиболее подходящих партнеров (половой отбор) и даже нередко ведут к половому дозреванию тех особей, которые в данный момент еще недостаточно подготовлены к размножению.
Перечисленные три канала брачной сигнализации часто применяются животными комплексно и, хотя все они были отнесены к дистантной рецепции дальнего действия, каждый из них имеет свои особенности и преимущества: звуковой канал — дистантный и хорошо применяется в условиях плохой видимости (заросли леса, водоросли водоема, темнота ночи), химический также может быть дистантным и, кроме того, следовой, т. е. надолго остающийся после ухода животного и, наконец, оптический позволяют с большой степенью точности определить местонахождение партнера, его внешние особенности и характер движений и мимики. На низких уровнях эволюции, пожалуй, преобладает химическая сигнализация, как это показано для коловраток, выделяющих в воду химические специфичные для вида вещества.
На более высоких ступенях эволюции, в ветви как первичноротых, так и вторичноротых к химической сигнализации подстраивается, все больше оптическая и акустическая сигнализация. Таким образом, как у млекопитающих (высшее звено ветви вторичноротых), так и у насекомых (высшее звено первичноротых) можно наблюдать обычно брачную сигнализацию, идущую по всем трем каналам. При этом у разных видов могут преобладать то один, то другие каналы. Так, например, самцы бабочки сатурнии (Saturnia pyri Schiff.) по запаху находили зрелых самок, прилетая к ним и ночной время на расстоянии 2 и более километров.
Пожалуй, больше всего внимания исследователи уделили брачным ритуалам у птиц. Дело в том, что в связи с уменьшенной возможностью у представителей этого класса к использованию хеморецепции у них особенно сильно развиты звуковая и зрительная сигнализации. Поэтому в данном случае исследователи располагают наилучшими возможностями регистрации сигналов, как с помощью зарисовок и фотокиноаппаратуры, так и средствами звукозаписи. В литературе описано очень много брачных сигналов и ритуалов у птиц, особенно у моногамов. Брачное ритуальное поведение у них имеет еще существенное значение в том отношении, что при этом будущие родители как бы демонстрируют друг другу свои способности к заботе о будущем потомстве. Как пример он приводит подражание самкой большой синицы и некоторых других птиц позе, движениям и голосу просящего пищу птенца, что вызывает реакцию самца на поиск насекомых и акт кормления. Поскольку в заботе о потомстве у птиц заключается важнейшая часть репродуктивного поведения, то именно по этой линии идет половой отбор при первоначальной встрече пар.
Песни воробьиных птиц имеют значение не только брачных церемоний ухаживания. Они обозначают также занятость данной территории самцом и его гнездом. При нарушении этой территории другими самцами они немедленно изгоняются. Вообще в поведении многих птиц в репродуктивный период у самцов обычно значительным элементом поведения становится агрессия, поскольку они должны охранять от других особей данного вида свое гнездо и прилежащий к нему кормовой участок, который необходим для обеспечения пищей будущего потомства. Это развито особенно у неколониальпых моногамов. В этом случае агрессия в период размножения имеет в первую очередь значение для занятия популяций вида более широкой площади и для обеспечения кормом развивающейся молодой генерации.
Самцы целого ряда видов птиц включают в ритуал привлечения самок обнаруженное ими место для гнезда. В качестве примера можно привести наблюдения за характерным поведением мухоловки-пеструшки [Muscicapa hypoleuca]. Обнаружив в середине мая пустую дуплянку, самец обследовал ее и начал усиленно петь и часто лазить в леток. Это продолжалось около полутора суток, причем самец пытался петь и около другой дуплянки, занятой большими синицами (Parus major), которые его активно гоняли. Через двое суток появилась самка мухоловка-пеструшка, которая заинтересовалась и самцом и его дуплянкой. Самец долго гонял самку (агрессивное поведение) и не давал ей залетать в дуплянку. Через 4—5 час он перестал ее гонять (в чем немалое значение имели ее умиротворяющие ритуальные движения) и допустил влезть в дуплянку, но только следом за собой. Еще через 2—3 часа самка начала носить в дуплянку материал для гнезда. При этом еще сутки самец пускал самку с материалом в дуплянку только после того, как он предварительно влетал туда и вылетал обратно. Лишь через сутки самка самостоятельно смогла беспрепятственно влетать в дуплянку. Можно было думать, что у самца угасло агрессивное поведение, но появившуюся еще через сутки вторую самку, которая тоже хотела осмотреть дуплянку, самец долго гонял и прогнал со своего участка. Таким образом, у наблюдателя создалось законное впечатление, что самец примерно через сутки научился узнавать именно свою самку и отличать ее от других. Агрессивность же его при защите своей гнездовой территории не угасла до момента вылета птенцов: любая залетевшая сюда «чужая» птица того же вида немедленно подвергалась нападению. Аналогичные наблюдения достаточно хорошо описаны в ряде статей и книг. Опыты подтверждают, что в защите гнездового участка нет права сильного, а есть право первого; хозяин, территории всегда победитель. Это же отметил А. Н. Промптов, указывая, что у воробьиных птиц при столкновениях из-за гнездового участка «победа, как правило, оказывается не на стороне сильного, а на стороне «законного владельца»; «агрессор», даже более сильный дерется слабее хозяина, отстаивающего свои права.
При неколониальном (одиночном) гнездовании птицы гораздо меньше защищены от врагов и поэтому тщательно прячут и маскируют свои гнезда, не отлетая далеко от них за пищей. Последнее заставляет их тщательно охранять от конкурентов пригнездовой кормовой участок. При этом, конечно, можно наблюдать все переходы от колониальных птиц (чаек, чистиковых и др.) к птицам слабоколониальным и одиночным. Например, дрозды-рябинники (Turdus pilaris) могут гнездиться и одиночно, но предпочитают делать гнезда небольшими колониями. В них гнезда располагаются довольно далеко одно от другого, но птицы защищают их от врагов все вместе. По тревожному крику одной птицы немедленно взлетают в воздух и собираются на крик все члены этой колонии. Поэтому возможные враги — вороны, сороки, сойки и белки — быстро научаются избегать территории, занятой группой гнезд дроздов-рябинников.
У птцц-полигамов, как правило, защита гнездовой территории отсутствует. У многих из них самцы весной собираются в определенные места — токовища, где поют брачные песни и устраивают турниры, основное значение которых — половой отбор. Самки, собираясь к токовищам, спариваются с наиболее крепкими и сильными самцами. Места токовищ обычно довольно постоянны, и как самцы, так и самки собираются сюда с довольно больших расстояний. У некоторых видов брачные песни на токовищах слышны лишь на очень небольшом расстоянии, как, например, у глухаря (Tetrao urogallus), или кавказского тетерева (Lyrurus mlokosiewiczi), в то время как у других, как, например, у тетерева-косача (Lyrurus tetrix), токовища слышны на несколько километров. Последнее может служить для ориентации при нахождении токовищ для самок и самцов. Это может иметь особое значение при меняющемся месте токовища, что нередко можно наблюдать именно у тетерева. В то же время такое дальнее звучание токовища может представлять собой и определенную опасность для популяции, поскольку на него могут ориентироваться и такие хищники, как ястреб-тетеревятник (Accipiter gentilis), а в утренних сумерках — u филин (Bubo bubo). Можно думать, что именно поэтому тетеревиные токовища обычно располагаются на безлесных пространствах с большой площадью обзора.
Для некоторых видов лягушек (остромордой Rana terrestris, озерной R. ridibunda, зеленой R. esculenta и др.) часто можно наблюдать брачные скопления во временных водоемах, которые «хоровым» пением ориентируют движение к этим водоемам и самцов u самок, пробуждающихся весной после зимнего анабиоза. Видимо, такие брачные скопления и их звуковая сигнализация и у амфибий и у птиц-полигамов являются определенными ориентирами для этих животных.
У большинства млекопитающих основным каналом брачной сигнализации является хеморецепция. Самцы узнают о готовности самок к размножению по издаваемым ими специфическим запахам. Конечно, у них большое значение имеют и звуковая сигнализация, и оптический канал, и осязание, широко дополняющие химическую сигнализацию. Хорошо известны ритуальные драки у самцов оленей и лосей, подбирающих себе гаремные стада. Известны ритуалы, сопутствующие образованию гаремов у таких ластоногих, как морские котики (Callorhinus ursinus) и сивучи (Eumetopias jubatus) . Они описаны рядом авторов и включают различные особенности совокупления, характерные для различных таксонов животных.
Забота о молодой генерации.
Следующим звеном репродуктивного поведения можно считать заботу о судьбе молодой нарождающейся генерации. Эту заботу, которая необходима для полноценного пополнения и существования популяции вида, можно видеть у животных, относящихся к ветви как первично-, так и вторичноротых. При этом можно проследить определенный прогресс в первой и во второй линиях.
Ветвь первичноротых.
Так, если у простейших, червей и большинства моллюсков не обнаруживается активной заботы о нарождающейся молоди, то у головоногих моллюсков можно наблюдать активную заботу и охрану отложенных кладок яиц. Так, например, у осьминога (Octopus) высоко развит материнский инстинкт: яйца, отложенные на донный субстрат, самка прикрывает своим телом, ни на минуту не покидая их и отказываясь от пищи длительное время. Во время этого «насиживания» самка часто омывает яйца струёй воды из воронки, промывая кладку. Многие самки осьминогов, закончив охрану кладки, погибают от истощения. Вылупившиеся же из яиц молодью особи головоногих моллюсков не охраняются.
В типе членистоногих (Arthropoda) для многих классов характерна активная забота о молоди, причем опять-таки в первую очередь охраняются яйцекладки. Так, например, панцирные клещи (Oribatidae) — чрезвычайно древняя группа животных — вынашивают яйца на своих панцирях вплоть до вылупления молоди. Лишь у некоторых паукообразных, ракообразных и насекомых можно видеть заботу о ювенильных особях. Многие пауки заботятся о своих яйцевых коконах и охраняют их. Самки пауков-волков (Lycosidae) при вылуплении молодых паучков держат кокон передними ногами, по которым они перебираются на спину матери, где живут некоторое время под ёе охраной. Долго продолжается забота о потомстве у паука-тенетника Theridium pictum. Молодые паучки этого вида совершено беспомощны и длительное время нуждаются в заботах матери Они живут на ее тенетах, питаются остатками ее стола и переходят к самостоятельной жизни, лишь достигнув значительных размеров. У паука Coelotes terreslris молодь остается в гнезде матери 34 дня после выхода из кокона За это время молодь трижды линяет. Мать защищает их от врагов. Она легко узнает их, переворачивая и ощупывая педипальпами. Паук других видов того же размера она убивает. Мать регулярно кормит свое потомство, давая им добычу, полупереваренную пищеварительными соками.
У большинства насекомых отсутствует активная забота о молодой генерации. Эта забота проявляется лишь в том, что самки откладывают яйца на тот субстрат, который необходим для питания личинок.
Многие жуки-навозники делают запасы навоза в специально выкопанных порах. На эти запасы навоза они откладывают яйца, обеспечивая, таким образом, пищей личинок, которые вылупятся из этих яиц.
Еще более сложной представляется забота о молодой генерации у представителей отряда перепончатокрылых (Hymenoptera), который вполне обоснованно считается наиболее высоко организованным в классе насекомых. Одиночные осы самых разных видов делают для своих будущих личинок соответствующие гнездышки. У одних видов они помещаются в норках, у других — в специальных домиках, напоминающих то маленькие горшочки, то другие гончарные изделия. В эти гнездышки самки одиночных ос складывают насекомых, парализованных (но не убитых) ловким уколом жала в определенные нервные центры. При этом осы разных видов запасают корма строго определенных видов: жуков-златок, долгоносиков, сверчков, кузнечиков, гусениц, некоторых пчел и даже пауков. В гнездо, в котором запасено достаточное количество парализованного корма, одинера сносит яичко, вход заделывает и улетает делать новое гнездо. Будущая личинка обеспечена пищей; больше о ней не заботятся. Значительно больше развита забота о потомстве у так называемых общественных перепончатокрылых — ос, шмелей, муравьев и термитов. Эти насекомые, живущие общинами, имеют в своих гнездах специальные камеры, или соты, для вывода и содержания личинок. Специальные «кормилицы», или «няньки», переносят яйца, личинок и куколок с одного места на другое, ориентируясь на температуру, влажность и чистоту камер. Все это многократно описано многими исследователями. Итак, п ветви первичноротых можно видеть, как постепенно в процессе эволюции развивается и усложняется забота о нарождающейся молодой генерации, что представляется весьма существенным для так называемой устойчивости популяции вида.
Ветвь вторичноротых.
То же можно проследить и в ветви вторичноротых, где забота о молодой генерации развивалась значительно более энергично. Так, у щетинкочелюстных (Chaetognatha), иглокожих (Echino-vermata) и оболочников (Tunicata) не наблюдается элементов активной заботы о молоди. У круглоротых (Cyclostomata) она сказывается лишь в закапывании икры в грунт. У надотряда костистых рыб (Teleoslei), объединяющего большинство видов современных рыб, можно видеть все переходы от видов, совершенно не имеющих активной заботы о потомстве, до видов с активной заботой об икре и молоди, держащейся в семейных группах.
Учитывая взаимоотношения родителей и потомства, класс рыб можно разделить на три группы: 1) производители покидают икру сразу после нереста; обучение в семейной группе отсутствует;
2) производители охраняют икру до выклева молоди; обучение в семейной группе отсутствует;
3) производители охраняют икру и образуют семейную группу с выклюнувшейся молодью; имеет место обучение в семейной группе.
Группа I. Рыбы - пелагофилы не заботятся ни об икре, ни о молоди. Они имеют обычно огромную плодовитость (луна-рыба Mola mola — 300 000 000, треска Gadus morhua — до 9 000 000 и т. д.) и соответственно ничтожный процент выживания.
Группа II. Некоторые виды рыб активно заботятся о выметанной икре. Икру, выметанную на субстрат, активно охраняют многие рыбы. Так, например, судак (Luciopеrсa luciopеrca L.) отметывает икру на прикорневую часть водных растении, после чего самцы, некоторое время издавая звуки угрозы (низкие, ударные), охраняют ее, защищая от врагов. Они отгоняют от кладок других рыб. Описаны случаи нападения судаков как на опущенную с лодки в воду pyку человека, так на подводных пловцов в аквалангах.
К этой же группе следует отнести рыб, строящих подобие гнезда и охраняющих там икру. К таковым в первую очередь можно отнести различных представителей семейства бычковых (Gobiidaе), особенно Азовского и Черного морей. Самцы этих рыб выкапывают норки в грунте или (что чаще) устраивают гнездовые пещерки под лежащими на дне камнями. Устроив такую пещерку, самец азовского бычка-кругляка особыми квакающими и скрипящими звуками подзывает готовых к нересту самок, которые выметывают икру, приклеивающуюся к стенкам и потолку пещерки. Окончив вымет, самки удаляются, а самец, оплодотворив икру, охраняет гнездо, издавая хриплые, отрывистые звуки, напоминающие рычание. Он не покидает свой пост при приближении опасности и набрасывается не только на крабов и судаков, но и на аквалангиста. Подобных наблюдений за рыбами семейства бычковых очень много.
Группа III. Ясно выраженная активная забота о своих мальках была установлена лишь для некоторых видов рыб тропических водоемов, где особенно велико напряжение отношений «хищник—жертва» и особенно значителен пресс хищников. Такое «семейное» поведение отмечено, прежде всего, у некоторых видов семейства хромисов (Cichlidae). Одни из них отметывают икру на грунт и ухаживают за ней, как представители группы II, другие вынашивают икру во рту, постоянно обмывая ее током проходящей воды. Однако и те, и другие образуют с активной молодью долговременные (до 30 дней) прочные семейные группы. При этом молодь стайкой плавает вместе с родителями, которые защищают ее от опасностей и обучают некоторым навыкам.
Таким образом, видно, как в классе рыб выработался новый тип поведения — активная забота о потомстве не только на стадии икры, но и на стадиях личинки и малька. У класса амфибий (Amphibia) в этом отношении наблюдается определенное отставание. Большинство представителей этого класса не заботятся активно ни об икре, ни о молоди. Лишь отдельные виды охраняют икру и заботятся о ней (например, углозубы — Нynobhdae, некоторые саламандры); самцы повитушки , (Alytes obstetricans) при спаривании наматывают икру на задние ноги и затем сидят с ней в ямке в земле; недели через две они уходят в водоем, где освобождаются от икры, из которой вскоре выходят головастики. У некоторых видов образуются приспособления для вынашивания икры до выклева личинок. Такова, например, южноамериканская суринамская жаба (Pipa americana), самка которой вынашивает икру и молодь в особых ячейках на коже спины. Вынашивает свою икру на спине в особом кожном кармане и самка сумчатой квакши (Notoderma). Можно привести еще ряд таких примеров. Однако ясно, что у амфибий дальше ухода за своей икрой забота о потомстве не развивается. Активной заботы о своей молоди и образовании семейных групп у них не бывает.
Почти то же можно сказать и о классе пресмыкающихся (Reptilia). Многие представители откладывают яйца в подходящий для них субстрат. Лишь некоторые змеи (питоны, кобры и американская иловая змея — Farancia abacura) охраняют кладку яиц до тех пор, пока вылупятся молодые змеи. Для самок американских аллигаторов (Alligator) известно, что они откладывают яйца в специально устроенные гнезда, выстланные растительными остатками, и охраняют их. Вылупившиеся молодые животные нередко собираются в так называемые «аллигаторовы ямы». Иногда молодые крокодилы держатся с матерью в течение многих месяцев.
По наблюдениям за крокодилами (Crocodylus moreleti) в зоопарке г. Атланта (США) самка продемонстрировала родительское поведение. Через 98 дней инкубации детеныши в отложенных яйцах стали издавать громкие звуки. Ориентируясь на эти звуки, самка вышла из воды и передними ногами раскопала кладку. После того как детеныши покинули скорлупу, самка осторожно, по одному детенышу брала их в пасть и относила к воде (на расстояние 2 м). Хотя мать и охраняла свой выводок, в течение двух месяцев все детеныши были съедены другими крокодилами.
Примерно те же особенности родительского поведения были отмечены и для нильского крокодила (Crocodylus niloticus). Самец и самка этого крокодила охраняют гнездо. После восприятия звуковых сигналов молодых особей самка раскапывает гнездо и переносит вылупившуюся молодь и яйца в пруд. Перекатывая яйца во рту, самка освобождает молодь от яйцевых оболочек.
У классов птиц (Aves) и млекопитающих (Manimalis) видно резкое, скачкообразное изменение поведения, направленного на |заботу о молодой нарождающейся генерации. Почти для всех видов этих классов характерна длительная забота о молоди, образование четко обусловленных семенных групп и довольно длительное (в разных группах по-разному) пребывание молодых животных вместе со своими родителями. «Родительское» поведение этих животных складывается из; следующих процессов: 1) нахождение места для гнезда или логова, 2) строительство гнезда для откладки яиц и пребывания ранней молоди, 3) насиживание яиц у птиц, 4) выкармливание и ухаживание за ранней молодью (птенцовые птицы), 5) выкармливание и обучение молоди, следующей за родителями семейными группами, 6) защита молоди от различных врагов.
Гнездостроение, так же как и выбор места для гнезда и защита находящихся там яиц и птенцов, имеет огромное значение для существования популяции данного вида. Именно период насиживания и выкорма молодых — наиболее опасное время для птиц, период наибольшей элиминации молодого поколения. Гибель этой молоди велика, причем, пожалуй, наибольшая — от различных хищников и других животных.
Строительство гнезд для высиживания яиц и выхаживания ранней молоди наиболее характерно для птиц. Лишь у некоторых видов не наблюдается специфического гнездостроения: одни откладывают яйца прямо на субстрат, как, например, кайры (Uria aalge) и некоторые кулички; другие вынашивают их на лапах, прикрывая складками кожи живота, как пингвины, некоторые подбрасывают яйца в гнезда других видов и, таким образом, являются «гнездовыми паразитами», например кукушки и некоторые другие виды. Многие виды сов, гнездящихся на деревьях, не строят своих гнезд. Tак, например, ушастая сова (Asio otus (L.)) иногда активно отбирает гнезда у сорок, ворон пли грачей., приспосабливая чужое гнездо, сова обычно почти не изменяет его конструкции. Огромное большинство видов птиц строит искусные гнезда, нередко поражающие своей сложностью и мастерством. Главное назначение этих сооружений — предохранение яиц и ранних птенцов (у птенцовых) от неблагоприятных факторов, прежде всего от резких изменений температуры, излишней влажности и различных хищников. Птицы каждого вида имеют свой стереотип устройства гнезда. И в то же время этот стереотип подвержен значительным изменениям, причем эти изменения поведения адекватны изменяющимся обстоятельствам.
Весьма интересно устройство гнезда некоторых птиц, например зяблика (Fringilla coelebs). Она представляет собой заключительный этап гнездостроения и является защитной маскировкой гнезда от глаз различных хищников. Поэтому, как правило, материалы для этой «конструкции» (кусочки лишайников) берутся с того ствола, на котором расположено гнездо.
У незрелорождающихся млекопитающих устройство гнезд обычно несколько проще, чем у птиц. Многие из них для рождения и вывода детенышей выкапывают в почве различной сложности норы, берлоги и логовища. Одни устраивают выводковые гнезда в дуплах деревьев, другие делают свои гнезда на растениях. Так, например, мышь-малютка прикрепляет свое шарообразное гнездо, свитое из тонких стеблей трав и растительного пуха, к стеблям злаков на расстоянии десятков сантиметров от земли. Гнездо белки, если оно не в дупле, также шарообразное, сделано изо мха, растрепанной коры деревьев u прутиков, помещается на хвойных деревьях обычно на значительной высоте. При этом часто самка белки строит несколько гнезд, в которые перетаскивает бельчат по мере заселения первого гнезда большим количеством паразитов. Гнездовое поведение представляет собой сложный комплекс врожденных актов: сбор материала, строительство гнезда, перенос детенышей и укрывание их подстилкой, обогрев, выкармливание молоком и защита гнезда.
Вскармливание потомства.
У птиц при вылуплении птенцов происходят новые резкие изменения поведения и активности птиц-родителей,происходит смена значений раздражителей в связи с переключением доминант периода насиживания и выкармливания. В период выкармливания птенцов происходят существенные перемены в общей активности взрослых птиц, так как воспитание потомства требует от родителей большой затраты сил.
После вылупления мощнейшим сигнальным фактором становятся именно птенцы. Гнездо теряет значение «ключевого раздражителя». При этом выводковые птицы (зрелорождающиеся) покидают гнездо в короткие сроки после вылупления, а птенцовые (незрелорождающиеся) остаются в нем довольно длительный срок. У первых почти сразу после вылупления четко действует рефлекс следования за самкой (или за обоими родителями), подчинение их сигналам (например, сбора, затаивания и т. д.), рефлекс подражания родителям и использования их для обогревания. Словом, у этой группы птиц сразу после выклева начинается активный контакт между родителями и потомством. На основе этого контакта образуются прочные семейные группы, примитивные формы которых существуют у рыб.
У вторых, рождающихся слепыми и беспомощными, первоначально действуют лишь безусловные рефлексы открывания рта и дефекации. Соответственно родительское поведение у этой группы птиц значительно сложнее, а контакт между родителями и потомством и прочные семейные группы здесь образуются и развиваются постепенно и сложно. Поскольку птенцовые птицы вылупляются из яиц слепыми, глухими и совершенно беспомощными, то их разделяют их ювенильный период жизни на три естественные стадии. Первой стадия - от момента вылупления до прозревания птенцов, когда последние еще лишены дистантной рецепции и преобладающими формами их поведения являются врожденные элементы. Второй стадией определяется период от включения зрительной и слуховой рецепции птенцов до оставления ими гнезда. В этот период у птенцов формируются многие начальные жизненно важные реакции. Наконец, третья стадия — от оставления гнезда до начала самостоятельной жизни. Для каждой стадии характерны особенности поведения как птенцов, так и родителей. Общей чертой всех стадий является кормление родителями птенцов. При этом резко возрастает активность сбора пищи при одновременном уменьшении ее потребления самими родителями. На первых двух стадиях происходит также уход за птенцами: чистка их покровов и вынос фекалия.
На всех трех стадиях родители охраняют и защищают птенцов самыми разными способами. Птицы ряда видов защищают потомство, самоотверженно и яростно нападая на приближающихся к гнезду врагов. Это характерно для многих хищных птиц, пускающих в ход когти и клювы. Чайки, крачки, а также дрозды-рябинники нападают на хищных птиц и зверей целой стаей и ударами с налета обычно обращают их в бегство. При нападении на приближающегося к гнезду человека они налетают на него и довольно точно попадают своими фекалиями. Чибисы около гнезд налетают на собак, лисиц и хищных птиц. Активно защищают своих птенцов (в гнездах) дрозды - белобровики в отличие от певчих и черных дроздов. Такая форма защиты потомства, характерна и для ряда рыб. Другая форма защиты заключается в затаивании самки на гнезде или около выводка и внезапном вылете на очень близком расстоянии от хищника (или человека). Часто при этом наблюдается так называемая «отвлекающая демонстрация. Она заключается в том, что птица так или иначе имитирует доступный для хищника пищевой объект, отвлекая его от гнезда или от выводка. Различают в этих отвлекающих демонстрациях несколько форм: 1) имитация убегающего зверька, когда птица бежит по земле с взъерошенными перьями спины, втянутой шеей и опущенным к земле хвостом; 2) имитация птенца с трепещущими крыльями и громким тонким ювенильным писком; 3) имитация раненой птицы с подлетами и падением на землю. Все эти формы известны и у других видов птиц, гнездящихся на земле или низко над землей. Так, например, хорошо известны отвлекающие демонстрации «раненой» птицы у самки рябчика около выводка. Элементы отвлекающего полета можно видеть у некоторых пеночек и славок.
В какой-то степени сходно с птицами проходит цикл репродуктивного поведения и у млекопитающих. У зрелорождающихся (например, копытные) родители не делают обычно гнезда или логова и детеныши уже через несколько часов после появления на свет могут следовать за матерью. У незрелорождающихся, как правило, имеет место устройство гнезда или логова с последующим выкармливанием детенышей сначала молоком, а затем обработанной пищей. Например, волк самец приносит выводку пищу (мясо), наполовину переваренную, которую он отрыгивает при соответствующих сигналах со стороны своего потомства.
У большинства млекопитающих родители (чаще всего самки) очень заботятся о своих детенышах, ухаживают за ними и самоотверженно защищают их. Таким образом, именно у млекопитающих особенно развит материнский инстинкт.
Потревоженные человеком и при других отрицательных моментах родители многих млекопитающих переносят свое потомство на новое место. Этот акт резко отличает млекопитающих от большинства птиц, у которых перетаскивание птенцов при опасности наблюдалось только у самки вальдшнепа.
Групповое поведение.
Часто можно наблюдать животных, относящихся к самым различным таксонам, которые образуют одновидовые концентрации особей и тем самым так или иначе вступают во взаимоотношения друг с другом и на короткий срок или длительно ведут групповой образ жизни.
Скопления.
Простейшее образование таких групп — называемых скоплениями, происходит не вследствие привлечения одних особей другими, а в результате привлечения в одно и то же место каждой особи независимо от других. Привлекающие факторы могут быть при этом самые разные: в одних случаях — большое количество пищи (мухи и другие насекомые у разлагающихся органических остатков, скопления травоядных животных на пастбищах, скопления у водопоев и т. п.), в других — тепловые и световые пятна (многие насекомые), многие запахи. В качестве примера приведем наблюдаемое нами скопление бабочек — тополевых ленточников (Limenitus populi) , которых привлек в одно место запах лисьего фекалия (наблюдение 17 топя 1972 г.)
Можно привести также пример образования скопления водомерок
(Hydrometra Sp.),
которые собрались в одно место (затишное от ветра) вблизи выступа берега на
одном из озер Киевской области. Как видно на рисунке, водомерки то держатся, не
мешая друг другу, довольно стройными рядами, то, будучи напуганными, собираются
в беспорядочное скопление. По окончании ветра они рассеиваются вдоль береговой
липни. 
Скопление водомерок, напуганное экспериментатором
Большие скопления образуются при миграциях животных — на местах отдыха, ночевки или у преград, возникающих на путях миграций (например, у мест переправ млекопитающих через реки).
Иначе обстоит дело в тех группах животных, которые создаются на более длительный срок в результате взаимного привлечения особей. Для них характерно, как правило, социальное, взаимообусловленное поведение, базирующееся на контактах и сложных комплексах сигналов.
Множество исследователей делят группы и групповое поведение животных на семейные и несемейные. Хотя семейные группы представляют собой лишь частный случай группового поведения, семейное групповое поведение имеет очень большое значение в жизни высших позвоночных животных.
Семейные группы.
Образование семейных групп рассматривается как следствие описанного выше репродуктивного поведения у большинства птиц и млекопитающих, а также у некоторых видов рыб и членистоногих. Эти группы состоят из родителей (или одного из них) и их потомства одной или даже нескольких генераций. Обычно родители доминируют в этих группах, кормят и защищают потомство и способствуют выработке у него необходимых для самостоятельной жизни реакций и навыков, что имеет очень большое значение в их экологии. Длительность существования семейных групп у животных разных классов и разных видов весьма различна: от нескольких декад до нескольких лет. Особенно длителен этот период у некоторых млекопитающих. Семейные группы и семейное поведение имеют место обычно до тех пор, пока молодые животные подрастут и приобретут основные черты поведения взрослых животных. Тогда происходит распад семейных групп, и животные переходят либо к одиночной жизни, либо к образованию несемейных групп (стад, стай).
Несемейное групповое поведение
Групповое поведение животных, относящихся к одному виду, представляет собой чрезвычайно интересное явление, поскольку оно, включает так или иначе проявляющуюся согласованность действий животных, входящих в данную группу. Эта согласованность обычно происходит на основе различной сигнализации и инстинкта подражания, которые позволяют группе действовать как единому целому. Д. В. Радаков отметил, что «групповая жизнь животных — это пример специфических закономерностей жизни, свойственных ее надорганизменному уровню и как таковая она представляет собой биологическую проблему большого теоретического значения». Действительно, групповое поведение и жизнь в группах (стаях, стадах, колониях), которое можно наблюдать у животных самых различных классов, следует рассматривать как адаптацию данной популяции, проявляющуюся на надорганизменном уровне. Именно так и рассматривается груповое (стайное) поведение животных: это поведение создает определенные преимущества всей группы в целом, давая ей возможность коллективно влиять на окружающую среду в сторону ее оптимизации. Поэтому групповой образ жизни возникает конвергентно у животных различных таксонов, и эти животные отличаются обычно высокой численностью и большой экологической устойчивостью.
Можно вслед за другими исследователями установить два основных типа несемейных группировок животных, которые различаются по особенностям взаимоотношении и поведения животных, составляющих группу:
1) с доминированием одних особей над другими (такие группы называются иерархическими);
2) стаи без доминирования. В первых взаимоотношения в группе и единство группы основаны на системе «господства» и «подчинения», что рассматривают как проявление разносторонности особей в области их поведения. Это значительно способствует существованию популяции вида в целом. Степень доминирования в стаях различных животных очень варьирует. Доминирующие особи часто сменяются. В одних случаях лидером является самец, в других — самка. Об иерархических группах и доминировании у животных существует большая литература, поскольку на это явление было обращено много внимания исследователей.
Стайное поведение.
 В группах второго типа иерархия и доминирование обычно отсутствуют.
Животные держатся вместе в силу инстинкта стайности. Если иерархические группы
можно наблюдать почти у всех классов позвоночных, то стаи без доминирования в
основном имеют место и особенно распространены в классе рыб. В какой-то
степени их можно предполагать в стаях воробьиных птиц. Однако наиболее
пристально они изучались именно в классе рыб. Дело в том, что стайные рыбы
представляют особую ценность в хозяйственном отношении. Кроме того, изучать
стайное поведение, механизмы этого поведения наиболее удобно на стаях рыб,
помещенных в аквариумы и бассейны, да и просто в водоемах с применением современной
техники (акустическая локация, авианаблюдения, подводные наблюдения и киносъемка).
Интенсивные исследования стайного поведения рыб проводил в лаборатории Д. В.
Радаков, который на основе своих работ написал интересную монографию
«Стайность рыб как экологическое явление» [1972]. В этой книге он дает свое
определение стаи рыб как «временной группы особей обычно одного вида, которые
находятся (все или большей частью) в одной фазе жизненного цикла, активно
поддерживают взаимный контакт и проявляют или могут проявлять в любой момент
организованность действий, биологически полезную, как правило, для всех особей
данной группы. Внешний облик стаи может часто и сильно изменяться в
зависимости от состояния рыб и условий, в которых они находятся».
В группах второго типа иерархия и доминирование обычно отсутствуют.
Животные держатся вместе в силу инстинкта стайности. Если иерархические группы
можно наблюдать почти у всех классов позвоночных, то стаи без доминирования в
основном имеют место и особенно распространены в классе рыб. В какой-то
степени их можно предполагать в стаях воробьиных птиц. Однако наиболее
пристально они изучались именно в классе рыб. Дело в том, что стайные рыбы
представляют особую ценность в хозяйственном отношении. Кроме того, изучать
стайное поведение, механизмы этого поведения наиболее удобно на стаях рыб,
помещенных в аквариумы и бассейны, да и просто в водоемах с применением современной
техники (акустическая локация, авианаблюдения, подводные наблюдения и киносъемка).
Интенсивные исследования стайного поведения рыб проводил в лаборатории Д. В.
Радаков, который на основе своих работ написал интересную монографию
«Стайность рыб как экологическое явление» [1972]. В этой книге он дает свое
определение стаи рыб как «временной группы особей обычно одного вида, которые
находятся (все или большей частью) в одной фазе жизненного цикла, активно
поддерживают взаимный контакт и проявляют или могут проявлять в любой момент
организованность действий, биологически полезную, как правило, для всех особей
данной группы. Внешний облик стаи может часто и сильно изменяться в
зависимости от состояния рыб и условий, в которых они находятся».
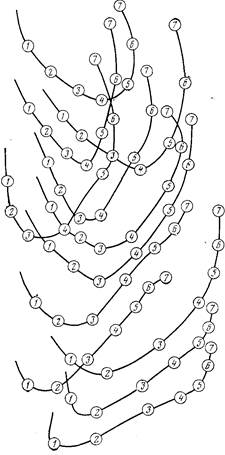
Основные типы структуры стаи пелагических рыб доказаны на схеме. Большое внимание Радаков уделил механизмам согласованности (или организованности) действий рыб в стае, что представляет интерес особенно в связи с отсутствием постоянных вожаков в стае рыб. В этом отношении стаю рыб, говоря языком кибернетики, следует рассматривать, как пример самоуправляемой системы без центрального управления. Опыты Радакова над некоторыми видами стайных рыб подтвердили вывод о том, что в стаях большинства рыб постоянные вожаки отсутствуют. При этом рыбы, идущие в головной части стаи, постоянно заменяются новыми из основной массы данной стаи. Расшифровка кадров киносъемки движущихся стай в экспериментальных бассейнах показала, как рыбы, движущиеся в головной части, даже при прямолинейном движении, постепенно отстают и оказываются в середине стаи, а при повороте на 180 градусов передние начинают поворот, но в поворот включаются все особи и в результате идущие в задней части оказываются впереди (см. рис.). Эти эксперименты также показали, что роль «вожака» на каждый данный момент выполняет достаточно большая по численности часть стаи. Так, для молоди сельди и карповых рыб было доказано, что изменение поведения и движения всей стаи определялось соответствующим изменением части стаи в том случае, если эта часть по своей численности составляла не менее 30— 40% от общего количества особей стаи. Сигнализация в данном случае заключается в передаче особенностей поведения и скорости движения определенной части стаи, выполняющей в этот момент функцию инициатора поведенческой реакции, остальным членам стаи.
Кроме того, экспериментируя в бассейнах Института океанологии Академии наук Республики Куба со стаями атериноморуса (Atherinomorus stipes Muller a. Troshel), Д. В. Радаков установил при помощи киносъемки, что в случае локального испуга у рыб, составляющих незначительную часть стаи, по всей стае пробегает «волна возбуждения». Это быстро перемещающаяся по стае сигнальная зона, в которой рыбы мгновенно реагируют на действия соседей измененном позы тела. Сами рыбы при этом почти не двигаются, но подгибают хвост, как бы готовясь к броску, а передвижение «волны возбуждения» достигает скорости 11,8— 15,1 м/сек, т. е. она в 10—15 раз превышает максимальную (бросковую) скорость плавания атериноморусов (рис. 28). Таким образом, сигнал испуга обычно передается по стае атериноморуса быстрее, чем за секунду. Далее этот сигнал может или затухнуть или вызовет «поток движения» всей стаи или ее части. «Поток движения» наблюдался в стаях почти всех исследованных видов рыб. В свою очередь, возникнув у части стаи, он может затухнуть или превратиться в «лавинообразный поток» всей стаи, что зависит от реактивности рыб, количества их в «потоке», скорости его движения и расстояния между «потоком» и остальными рыбами стаи. В большой степени общая реакция стаи зависит и от силы и направления пугающего раздражителя.
Защитное значение стаи.
Для животных, находящихся в естественных условиях, там, где они, как правило, окружены врагами, скопление в многочисленные группы, казалось бы, должно было увеличить их способность к обороне, если сами эти группы не имеют оборонительных способностей. Но поскольку в группах (стаях, стадах, колониях) держатся (по временам или постоянно) животные, относящиеся к весьма различным таксонам, невольно приходит мысль о том, что именно такие группы представляют собой конвергентные оборонительные адаптации, служащие для сохранения численности популяции вида.
И, действительно, исследования выявляют все больший «арсенал» оборонительных возможностей организованной группы животных. Прежде всего, группа животных, которая ведет «тактику кругового обзора», замечает своего врага на значительно большем расстоянии, чем одна особь. Поэтому хищнику намного труднее приблизиться к группе животных на расстояние броска. Одиночные гольяны легче становились добычей щуки. В стаях большинства позвоночных животные могут более спокойно отдыхать или питаться, поскольку часть из них (случайно или даже специально) выполняет роль «часовых» и при появлении опасности движениями или звуками настораживает всю группу. Затем следуют различные оборонительные действия всей группы.
Животные ряда видов, объединившись в группы, активно обороняются от врагов и даже нападают на них. Такое поведение известно для копытных животных (быков, вилорогов и овцебыков). Эти животные при нападении волков и некоторых других хищников часто образуют каре, и, спрятав телят в середину, становятся рогами наружу, организуя круговую защиту. Морские чайки, так же как и вороны, объединившись в гнездовые колонии, часто нападают на хищников и прогоняют их. Следует вспомнить, что активные способы групповой защиты существуют и в ветви первичноротых, где ряд видов общественных перепончатокрылых активно защищают свои гнезда и колонии коллективно, нападая на врагов и пуская в ход свое «оружие».
Такая активная защита — нападение характерна для тех животных, ведущих групповой образ жизни, которые по тем или иным причинам не могут спасаться от врагов бегством, будучи приуроченными к постоянным местам (гнезда с потомством, колонии перепончатокрылых, слабый молодняк) и в то же время имеют различные возможности нападения.
Многие стайные животные спасаются от хищников, убегая, улетая или уплывая от них тесной группой. Казалось бы, увеличенное количество особей в стае увеличивает возможность их поимки хищником, но данные научных исследований показывают обратное: в ряде случаев рыбы, птицы и млекопитающие, так же как и некоторые другие животные, держась в стаях, оказываются менее доступными или даже совсем недоступными для хищников. Даже рыбы, питаясь беспозвоночными (например, дафниями) , находящимися в плотных скоплениях, поедают их менее интенсивно, чем при более разреженных концентрациях. Такое явление называют «эффектом смущения» хищника многочисленностью жертв. В погоне за стаей рыб дневной хищник как бы «дезориентируется» большим количеством мелькающих рыбешек, его погоня становится менее целеустремленной, броски следуют один за другим и их подавляющее большинство оканчивается промахами. В то же время погоня за одной рыбкой проходит очень направленно и завершается одним удачным броском». Это дало основание назвать описываемое явление «дезориентацией хищника» вследствие многочисленности жертв.
Дезориентация хищника еще более увеличивается в результате особых защитных «маневров» стаи. Эти маневры неоднократно наблюдал и фиксировал Д. В. Радаков киносъемкой для целого ряда морских и пресноводных рыб, как в отношении живых хищников, так и их моделей. «Маневрирование» заключается в том, что при броске хищника на стаю, находящуюся в состоянии кругового обзора, рыбы ближайшей части стаи рассыпаются от хищника веером вперед и в стороны, создавая перед мордой хищника постоянную «пустоту», и, немного проплывая, тут же заворачивают к хвосту хищника против направления его броска. При этом часто стая, разделившись на две части, снова соединяется и следит за удаляющимся хищником. Этот маневр, если его нанести на бумагу, похож на букву Ф, причем путь хищника составляет вертикальную часть этой буквы (см. рис.А). За это сходство называют такой маневр стаи условно «Ф-маневром». Такое маневрирование было зафиксировано для целого ряда рыб в экспериментах в больших бассейнах. Они были отмечены при погоне кефали и морского налима за стаями атерины (Atherina mochon pontica Euch.), саргана (Belone belone (L.)) за стаями хамсы (Engraulis encrasicholus (L.)), ставриды (Trachurus mediterraneus ponticus Aleev), за стаями мальков кефали, щуки, за стаями верховок и в ряде других случаев.
для стаи песчанки (Ammodytidae), преследуемой пловцом. В момент внезапного испуга (например, бросок хищника) стая мелких рыб часто рассыпается веером, что также дезориентирует хищника. Рассеявшаяся таким образом стая обычно снова быстро восстанавливается. Следует заметить, что картина реагирования стаи пелагических рыб на хищника и специфика ее маневрирования в значительной степени зависят также от соотношения направлении движения стаи и движения хищника.
Указанные особенности стайного поведения рыб в условиях дневной освещенности значительно затрудняют охоту хищников за рыбами, находящимися в стае. Эксперименты, проведенные Д. В. Радаковым и его сотрудниками, показали примерно одно и то же: рыбы в стаях при нападении хищников оказывались для них значительно менее доступными, чем одиночные особи, и истреблялись в 5—6 раз медленнее. Это было доказано как на морских, так и на пресноводных рыбах. Как пишет Радаков, «хищник, нападая на стаю, не преследует какой-нибудь одной рыбки до тех пор, пока не поймает ее. Погнавшись сначала за одной и упустив се, он устремляется за другой, за третьей, пока, наконец, ему удается схватить одну из жертв. В результате, на ее поимку уходит больше времени, чем, если в аквариуме находится одиночная рыбка, погоня за которой получается более целеустремленной».
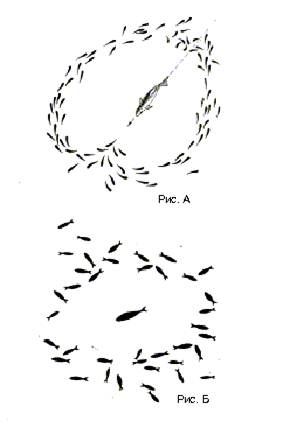 Обычно
голодные хищники, помещенные при достаточной освещенности вместе со стайкой
рыб-жертв, в первые минуты начинали энергичную погоню и за это время иногда
успевали схватить несколько рыбок. За эти первые минуты в результате пугающего
воздействия хищника стая уплотнялась, принимая «оборонительную» структуру (см.
рис. Б). Это в еще большей степени снижало эффективность охоты, соответственно
его пищевая активность уменьшалась, а в ряде случаев прекращалась совсем. Можно
считать, что прекращение охоты связано с том, что энергия, затрачиваемая
хищником на погоню, оказывается значительно большей, чем энергия, получаемая от
пищи. Таким образом, охота становится энергетически невыгодной.
Обычно
голодные хищники, помещенные при достаточной освещенности вместе со стайкой
рыб-жертв, в первые минуты начинали энергичную погоню и за это время иногда
успевали схватить несколько рыбок. За эти первые минуты в результате пугающего
воздействия хищника стая уплотнялась, принимая «оборонительную» структуру (см.
рис. Б). Это в еще большей степени снижало эффективность охоты, соответственно
его пищевая активность уменьшалась, а в ряде случаев прекращалась совсем. Можно
считать, что прекращение охоты связано с том, что энергия, затрачиваемая
хищником на погоню, оказывается значительно большей, чем энергия, получаемая от
пищи. Таким образом, охота становится энергетически невыгодной.
При изучении оборонительного значения стайного поведения рыб особый интерес представляет их химическая защитная сигнализация. Впервые эту сигнализацию обнаружил Фриш [Frisch, 1938], установивший, что при ранении одного гольяна вся стайка становилась испуганной, разбегалась или уходила в сторону. Фриш показал, что такое же действие оказывает на стаю экстракт из кожи только что убитого гольяна. Эти исследования, продолженные Фришем и другими исследователями, показали, что в коже целого ряда видов рыб находятся специальные колбовидные клетки, не имеющие связи с поверхностью и содержащие вещества, которые при ранении кожи выходят в воду и немедленно вызывают у рыб данного вида сильнейшую реакцию испуга. Это вещество названо «веществом испуга», и установлено, что оно воспринимается при помощи обоняния даже в очень небольших концентрациях. Фриш в опытах с гольянами подсчитал, что пороговая концентрация этого вещества в воде равнялась примерно 1,4 •1010 г/л. «Вещество испуга» (иногда его называют «феромоном тревоги») и соответствующие реакции были обнаружены у подавляющего большинства рыб отряда карпообразных (Cyprinifornies) и у некоторых видов из других отрядов. Это действие у рыб разных экологических групп проходило по-разному: рыбы, обитающие в зарослях и у других укрытий, состаивались и четко ориентировались на источник запаха, а затем затаивались или уходили в укрытия; придонные рыбы после кратковременного состаивания и броска от источника запаха затаивались у дна на длительное время; рыбы, обитающие в толще воды и у поверхности, реагировали уходом или бросками, а затем снижали активность с образованием плотной защитной стайки. Таким образом, можно сделать вывод, что при воздействии «вещества испуга» образуются определенные экологические стереотипы оборонительного поведения рыб.
Весьма близко к указанной сигнализации стоит явление «запаха страха», установленное для грызунов. Запах, оставленный раненой живой домовой мышью, отпугивает от данного места се сородичей. Было отмечено, что поскольку пятна крови и остатки шерсти мышью не оказывают такого отпугивающего действия на других мышей, то можно думать, что «запах страха» выделяется соответствующими железами испуганного зверька. Наличие таких сигналов, приносящих пользу всей стае, или популяции, еще раз подчеркивает правоту вывода Радакова о том, что групповая жизнь животных, и в частности стайное поведение,— это явление, характерное для надорганизменного уровня, это — групповая защитная адаптация, которая могла создаться в результате группового, а не индивидуального отбора.
Защитное значение стаи известно и для ряда птиц. Ориентолог В. Э. Якоби [1967] пишет, что плотные и быстроманеврирующие стаи скворцов, а также некоторых болотных птиц мешают хищникам, и в частности соколу-сапсану, прицельно и успешно атаковать и схватить определенную птицу. Поэтому хищные птицы при нападении на стаю, прежде всего, стараются отбить от стаи одну особь, а потом уже хватают ее. Часто же при нападении ястреба на стаю мелких птиц он не может схватить ни одной из них.
Для некоторых копытных животных стайное скопление имеет определенное защитное значение также по отношению к кровососущим насекомым. Летом при обилии гнуса (оводов, комаров, кровососущих мух) северные олени собираются в плотное стадо. Кровососы обычно облепляют оленей, находящихся в наружных рядах, и почти не проникают в глубь стада. Поэтому животные в центре стада спокойно стоят или лежат, а внешние ряды оленей ведут себя беспокойно и постепенно передвигаются вокруг центра стада. Чем активнее ведут себя кровососы, тем больше внешних рядов стада оленей находятся в движении, но их число обычно не превышает пяти. Время от времени крайние олени, измученные гнусом, силой прорываются в центр, расталкивая соседей. Учитывая количество оленей в стаде и количество оленей во внешних (беспокойных) рядах, подсчитано, что при 500 оленях в стаде, защищено от кровососов 56% стада, при 2000—77%, а при 4000-83%.
 Говоря о защитном значении группового поведения, следует
отметить и защиту животных от неблагоприятных абиотических факторов среды. В
целом ряде работ можно найти данные о том, что животные, собравшись в группу,
тем самым как бы влияют здесь на микроклимат и потому легче переносят ветры,
метели, чрезмерную низкую или высокую температуру. Взаимное обогревание и коллективную
регуляцию температуры в группах животных самых разных таксонов отмечали
большое количество исследователей. Оно известно и в колониях общественных
насекомых (пчелы, муравьи), и при ночевках некоторых птиц и для ряда стадных
млекопитающих. Неоднократно описаны скопления пингвинов во время морозных
ураганов. Эти антарктические птицы образуют плотные тысячные стаи, в которых
птицы со стороны постепенно передвигаются на подветренную. При этом огромная их
масса постоянно «ползет», подгоняемая ветром. Такое движущееся скопление
пингвинов иногда называют «черепахой». Сходно ведут себя во время снежных
буранов стада овец, лошадей, антилоп и северных оленей. В степях и пустынях в
летние жаркие дни овцы также образуют скопления, пряча свои головы в тени,
отбрасываемой сочленами стада. Наконец, многие рыбы, змеи и некоторые
млекопитающие, впадая в зимнюю спячку, также образуют большие зимовочные скопления,
в которых значительно снижается уровень обмена веществ.
Говоря о защитном значении группового поведения, следует
отметить и защиту животных от неблагоприятных абиотических факторов среды. В
целом ряде работ можно найти данные о том, что животные, собравшись в группу,
тем самым как бы влияют здесь на микроклимат и потому легче переносят ветры,
метели, чрезмерную низкую или высокую температуру. Взаимное обогревание и коллективную
регуляцию температуры в группах животных самых разных таксонов отмечали
большое количество исследователей. Оно известно и в колониях общественных
насекомых (пчелы, муравьи), и при ночевках некоторых птиц и для ряда стадных
млекопитающих. Неоднократно описаны скопления пингвинов во время морозных
ураганов. Эти антарктические птицы образуют плотные тысячные стаи, в которых
птицы со стороны постепенно передвигаются на подветренную. При этом огромная их
масса постоянно «ползет», подгоняемая ветром. Такое движущееся скопление
пингвинов иногда называют «черепахой». Сходно ведут себя во время снежных
буранов стада овец, лошадей, антилоп и северных оленей. В степях и пустынях в
летние жаркие дни овцы также образуют скопления, пряча свои головы в тени,
отбрасываемой сочленами стада. Наконец, многие рыбы, змеи и некоторые
млекопитающие, впадая в зимнюю спячку, также образуют большие зимовочные скопления,
в которых значительно снижается уровень обмена веществ.
Значение стаи при питании.
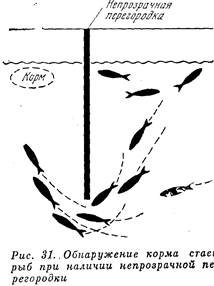
Значение стаи (пли вообще группирования) животных при питании также довольно разнообразно. Прежде всего, в группах животные легче находят скопления пищи. Как показали опыты, проведенные с молодью сайды, та часть рыб из стаи, которая обнаружила корм и бросалась к нему, увлекала за собой других рыб стаи, которые не могли видеть корма (он был от этих рыб скрыт непрозрачной перегородкой), а те в свою очередь увлекали еще более отдаленных сочленов стаи (См. рис3.1) . Таким образом, стадность облегчала рыбе поиск пищи, и в считанные секунды вся стая собиралась на скоплении кормовых организмов, обнаруженных лишь частью ее сочленов.
Велико значение стаи и при поимке добычи у тех хищников, которые применяют тактику «коллективной охоты». Выше было показано, что рыбы, держащиеся оборонительными стаями, становятся почти недоступными для одиночных хищников. Однако в качестве коадаптации у некоторых хищников выработался стайный способ охоты за стайными жертвами. Крупные окуни стаей окружают стайку молоди карповых рыб, отгоняют ее от укрытий и поедают Примерно также описано подобное явление для хищных рыб тропических морей. Д. В. Радаков приводит два своих наблюдения: днем у Западной Африки на поверхности воды, было замечено несколько стай анчоусов, преследуемых снизу тупцами и акулами, а сверху серыми буревестниками. Над стаями стояла пена и брызги. Стаи имели около 5 м в диаметре. Вскоре стаи были уничтожены, и на их месте можно было видеть лишь медленно погружающуюся чешую. Второе наблюдение было произведено в Черном море около Карадагской биологической станции, где Д. В. Радакову удалось приблизиться в подводной маске к стае ставрид, напавших на стаю песчанки. Песчанка держалась в очень плотной стае около полуметра в поперечнике и преследуемая снизу ставридами, была «буквально прижата к поверхности поды». Численность этой стайки быстро уменьшалась. Исходя из этих наблюдений, Д. В. Радаков заключает, что стая хищных рыб прижимает стаю своей добычи к поверхности воды снизу, в результате чего рыбы этой стаи не могут ни спастись бегством в стороны, ни скрыться в глубину. Далее этот автор делает общий вывод, что стайное поведение хищных рыб представляет собой адаптацию, способствующую поимке добычи, поскольку стая хищников может:
1) легче обнаружить стаю жертв и приблизиться к ней;
2) окружить добычу, препятствуя ее бегству;
3) оттеснить добычу от обычных укрытий и, в частности, «прижать» ее снизу к поверхности воды;
4) дезориентировать стаю жертв и внести в ее поведение элементы паники. Таким образом, стайное, организованное поведение рыб хищников оказывается полезным для всей группы в отношении питания. Это верно именно для стай, отличающихся взаимообусловленным, согласованным поведением, тогда как для простого скопления индивидуумов без согласованного поведения вполне подходит заключение «чем больше ртов, тем меньше на долю каждого».
Широко известна «коллективная» стайная охота хищников из семейства собачьих, при которой употребляются самые разные приемы: «оцепление», «гоньба», «нагон», «подстанавливание» и т. п. Они описаны для волков, гиеновых собак, австралийских динго и некоторых других хищников. Коллективная охота описана и для касаток. Эти китообразные охотятся всегда стадом, причем при охоте и на моржей и на дельфинов их приемы были сходны: «сначала следовало окружение стада, а затем расправа с жертвами.
Значение стаи при миграциях и размножении.
Большая часть мигрирующих животных совершает миграции, собираясь в большие стаи, объединяющиеся в передвигающиеся скопления. Исходя из этого, можно полагать, что групповое поведение представляет собой важную адаптацию и при миграциях животных. По всей вероятности, стайность и групповое поведение и в этом случае важны, прежде всего, в защитном и пищевом отношениях. Для животных, передвигающихся по неосвоеннным ими пространствам, защита от врагов и обнаружение мест скопления пищи и мест отдыха должны иметь первоочередное значение. Возможно, в стаях животные легче ориентируются при миграциях. Наконец, весьма вероятно, что стайные миграции рыб имеют прямое отношение к гидродинамическим расчетам, которые показали, что стая рыб, плывущая определенным строем, затрачивает значительно меньшую энергию. В общем же следует заметить, что значение стайного поведения животных при миграциях изучено совершенно недостаточно и нуждается в дальнейших исследованиях.
Еще меньше изучено значение группового поведения животных при размножении. Некоторые позвоночные образуют в этот период скопления типа гнездовых колоний (у птиц и рыб) или лежбищ (у ластоногих). Многие рыбы, подходя к нерестилищу большими скоплениями стай, размножаются, продолжая оставаться в этих скоплениях. Так, например, баренцевоморская треска нерестует у берегов Норвегии, собравшись в большие косяки. Измеренный при помощи эхолота нерестовый косяк имел протяжение и ширину более километра, а толщина его составляла 10—15 м. Такое скопление состояло, по произведенным подсчетам, из нескольких миллионов особей
Следует отметить, что массовые скопления при размножении отмечены и у некоторых беспозвоночных. Так, неоднократно описаны подъемы от дна к поверхности моря нереид, которые временами образуют у поверхности огромные скопления. Интересный случай произошёл летом 1944 г на Белом море, вблизи берегов внезапно появилась масса нереисов (Nereis virens). Они плавали на поверхности моря, изгибаясь, как змеи. Тела их были длиной 30—40 см. В тихую погоду вода буквально кишела этими животными. Рыбаки даже были вынуждены прекратить лов рыбы и сообщить, что на море появились «морские змеи». Обычно эти черви живут на дне, а когда начинают созревать половые продукты, всплывают к поверхности воды для размножения. Тысячи нереид внезапно появляются в воде и «роятся» — плавают, змеевидно изгибаясь, до тех пор, пока половые продукты не выйдут в воду.
Можно предполагать, что все указанные группировки и скопления животных также полифункциональные и могут иметь значение как для интенсификации и синхронизации процессов размножения, так и для защиты от уничтожения хищниками производителей. Возможно также, что собравшиеся животные вносят свою молодую генерацию в больших концентрациях в наиболее оптимальные для нее условия.
Непостоянство стайности.
. Следует еще упомянуть об относительном непостоянстве, изменчивости несемейных групп группового поведения животных. У многих видов животных группы (стаи,. стада) образуются лишь на определенных этапах жизненного цикла (миграции, зимовки н т. д.), а при размножении они распадаются на пары и семейные группы. Так обстоит дело у многих птиц и у некоторых рыб. Кроме того, у образовавшихся стай очень часто меняется их состав в результате перемешивания. Так что нельзя со всей уверенностью сказать, что группы – явление постоянное.
Опосредованное обучение.
Почти у всех видов млекопитающих и птиц, а также у многих видов рыб имеет место явление, которое называют опосредованным обучением: это взаимообучение животных, приобретение ими при общении новых элементов поведения, повышающих устойчивость, «надежность» популяции в борьбе за существование. Опосредованное обучение обычно происходит на основе врожденной способности животных к подражанию, часто подкрепляется определенной сигнализацией и закрепляется памятью. Поэтому условные рефлексы, приобретаемые в результате опосредованного обучения, физиологи нередко называют условными имитационными рефлексами.
Хотя сам факт опосредованного обучения у позвоночных общеизвестен, он подтвержден экспериментальными данными еще весьма недостаточно.
Эксперименты над млекопитающими четко доказали, что животные исключительно легко образуют условно-рефлекторные связи в условиях общения. Условные рефлексы при общении животных вырабатываются в 2—3 раза быстрее, чем в одиночестве, что связано с подражанием.
Самое главное значение опосредованного обучения заключается в том, что при этом необходимый для жизни животного в тех или иных условиях внешней среды набор приобретаемых элементов поведения создается без непосредственного воздействия на данное животное безусловного раздражителя. Это очень важно при жизни в естественных условиях. Животное получает, например, оборонительные реакции к данному хищнику, не испытав на себе его прямого воздействия, без болевого раздражителя. Ему достаточно лишь видеть, как хищник схватил их соседа по группе, а в ряде случаев наблюдать, как соседи избегают то или иное опасное животное, или просто воспринимать их сигналы опасности. То же можно сказать и в отношении ядовитой пищи:
необязательно животным самим испытывать ее ядовитое действие. Вполне достаточно понаблюдать, что едят или чего избегают его более «опытные» соседи. При получении опыта индивидуальным путем каждое животное должно на себе непосредственно испытать вредное действие хищника или ядовитой пищи, что обычно в природе влечет за собой гибель животного. В группе этот опасный безусловный раздражитель заменен примером более опытных особей, а также их сигналами, передающими информацию о надвигающейся опасности. Таким путем необходимые особенности поведения (опыт) передается в общении с себе подобными в естественных условиях с неизмеримо меньшим числом ошибок, а, следовательно, и с меньшей потерей численности популяции. В этом, заключается прежде всего огромное адаптивное значение опосредованного обучения. Имитационное поведение — это главный охранитель вида, так как громадное преимущество заключается в том, что «зрители», присутствующие при акте повреждения члена их же стада пли их сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности.
Американский натуралист - Э. Сетон - Томпсон пишет, что у каждого дикого животного есть три источника познания. Первый источник — это опыт предков, инстинкт, переданный ему по наследству. Этот опыт накопился у целого ряда поколений за долгие века борьбы с опасностями. Второй источник познания — пример родителей и других взрослых животных того же вида. Детеныш перенимает все обычаи и повадки своего племени. Третий источник познания — собственный опыт.
Наследственный инстинкт не всегда помогает животному, так как он недостаточно изменчив и подвижен, а условия жизни постоянно меняются. Пример взрослых тоже не может научить детеныша всему необходимому. А третий источник знания плох тем, что личный опыт всегда приобретается слишком опасным путем.
Конечно, написанное относится в основном лишь к млекопитающим и птицам. Можно думать, что Э. Сетон Томпсон приуменьшает при этом значение опосредованного обучения, но в основном с ним следует согласиться.
Можно говорить о двух типах опосредованного обучения, постоянно переплетающихся и дополняющих друг друга: обучение в несемейных группах животных и обучение в семейных группах.
Групповое обучение в несемейных группах.
Обучение в несемейных группах (кратко — групповое обучение) происходит в стаях и стадах животных, в их временных скоплениях, в их территориальных группах и сообществах. Значение этих групп, и в частности стай, весьма многогранно. Пожалуй, лучше всего оно изучено на рыбах. Еще в опытах было Д. В. Радакова показано, что золотые рыбки (Caras-sius auratus (L.)) научаются проплывать лабиринт значительно быстрее, если они перед этим видели, как это делают другие, уже обученные рыбы.
Интересный опыт был проведен с карповыми рыбами. Разделив аквариум на две части прозрачной перегородкой, исследователи вырабатывали у рыб (карасей и карпов), помещенных в одном отсеке, условные пищевые двигательные рефлексы (положительный на зеленый свет и дифференцировочный—на красный свет). Условные рефлексы у этих рыб, которых назвали «актерами», проявились и закрепились достаточно быстро. Но самое интересное, что у другой группы рыб, названной «зрителями», которые никакого пищевого подкрепления не получали и могли лишь видеть, как его за стеклом получали «актеры», также выработались довольно прочные положительные условные рефлексы на зеленый свет. Эти рефлексы во времени имели волнообразный характер и затухали примерно на девятом-десятом повторении. Представляет интерес и тот момент, что в том случае, если «зрители» видели только выработку условного рефлекса (хватание бусинки) у «актеров», но лишены были возможности видеть акт пищевого подкрепления (он производился в данном случае за непрозрачной перегородкой), то выработать у них «подражательный» рефлекс не удавалось.
В августе 1978 г. в старице Днепра плавала стая язей длиной 16—17 см. Они были очень активны и явно голодны. На удочку, наживленную кузнечиком, бросилась сразу вся стая. Один, схвативший наживку, был вытащен. На брошенную вторично наживку они бросились не сразу, а минут через 5. Третий взял наживку через 15 мин, а затем на протяжении часа ее больше не схватил ни один. Все подходили к наживке и испуганно уплывали прочь. Следовательно, у них оборонительный рефлекс на удочку создался лишь при виде вытаскиваемых из воды сочленов стаи.
Все эти эксперименты достаточно четко доказывают, что у рыб, находящихся в стае, хорошо образуются как пищевые, так и оборонительные условные рефлексы, что свидетельствует об их способностям к групповому опосредованному обучению.
Можно думать, что в стаях птиц и млекопитающих происходит такое же групповое опосредованное обучение.
В литературе описано много случаев, когда молодые воробьи, вливаясь в стаи несколько более старших воробьев (предыдущего выводка), быстрее «приспосабливаются к условиям существования». Отмечено, что подражание в форме «актера» и «зрителя» широко используется разными видами млекопитающих при образовании новых условных связей. В частности, он наблюдал это у лабораторных мышей, причем «зрители» образовывали условный рефлекс на раздражитель, подкрепляемый актом еды «актера». Об обучении в стадах копытных много фактов приведено Л. М. Баскиным который устанавливает, например, что «формирование поведения оленей, находящихся в стаде, происходит в значительной мере за счет подражания». В отношении одомашненных животных отмечаено, что «если теленка, умеющего поедать траву, сено и другие корма, поместить в группу сверстников, не способных это делать, то они, подражая своему «опытному сородичу», через несколько дней приобретают те же навыки». И далее: «При появлении в стаде, отаре, табуне одного-двух животных, обладающих способностью преодолевать изгороди, рвы, водные преграды, большинство других в короткий срок научились приемам своих менторов».
У дельфинов превосходно развит рефлекс подражания. Если один дельфин в стайке научился, к примеру, пригибать сеть и таким образом выходить из окружения, этот опыт тут же перенимают другие. Такое подражание значительно облегчает дрессировку дельфинов и сокращает сроки их обучения. Те дельфины, которые долго находились с дрессируемыми сородичами, могли выполнять это действие даже без специальной тренировки. В Калифорнийском океанариуме малая косатка обучилась трюкам только на основе наблюдения за своими компаньонами — афалинами, с которыми работали тренеры.
Исходя из изложенного, можно говорить о том, что в группе животных открывается возможность использования опыта немногих особей всей группой.
Опосредованное обучение в семейных группах.
Это интереснейшее явление — обучение молодых животных их родителями — наиболее сильно развито у птиц и млекопитающих. На него обратил серьезное внимание Д. В. Радаков, назвав его термином «сигнальная наследственность». Это явление широко известно, но еще весьма недостаточно исследовано. Оно происходит в результате так называемого биологического контакта поколений и представляет собой чисто функциональную преемственность приспособительных реакций. При этом предшествующие поколения путем научения передают последующим поколениям накопленную ими информацию и соответствующие особенности поведения. Сами эта. особенности не врожденные, не закреплены генетически, но настойчиво передаются потомству в силу наследственного рефлекса подражания родителям или при помощи специальной сигнализации. При этом происходит как бы передача «по наследству» ненаследственной информации, не входящей в генетическую структуру организма, по через подражание и память настойчиво передающейся из поколения в поколение и в то же время постоянно изменяющейся адекватно изменениям факторов среды с каждым последующим поколением.
Способность к сигнальной преемственности, широко развитая у высших позвоночных и связанная, конечно, с прогрессивным развитием их нервной системы, чрезвычайно обогатила поведенческие, адаптивные особенности этих животных. Сигнальная преемственность стала здесь как бы дополнительным звеном между врожденными элементами поведения, относительно стабильными, и индивидуально приобретаемыми элементами, чрезвычайно лабильными. Она значительно обогатила и усовершенствовала поведенческий комплекс животных, объединив опыт многих поколений и способствуя образованию у них разнообразной и сложной сигнализации.
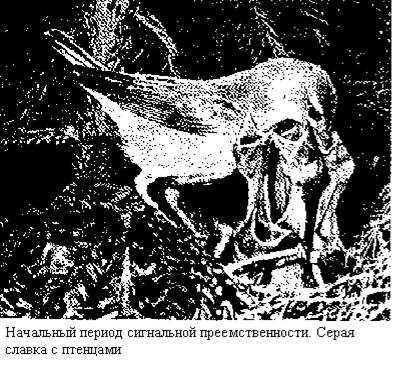
Мы полагаем, что в начале сигнальной преемственности стоит явление запечатлевания, установленное для многих видов позвоночных животных (главным образом для птиц).
У животных многих видов в онтогенезе имеются определенные «чувствительные периоды», во время которых они научаются реагировать на определенный, строго специфический комплекс раздражителей. Новорожденное животное, когда у него начинают действовать рецепторные системы, запоминает (запечатлевает) в короткий срок и нередко па длительный период окружающую его обстановку и прежде всего своих родителей — их внешний вид, голос, а у млекопитающих — и запах. Конрад Лоренц ввел термин запечатлевания для обозначения уникального процесса обучения, когда на ранних стадиях развития выводковые птицы образуют привязанность к внешнему виду матери. Именно запечатлевание родителей и стремление на какой-то срок им подчиняться и подражать создают прочную силу закона для сигнальной преемственности.
С физиологической точки зрения запечатлевание подробно разобрано. Многие этологи проводили эксперименты, показывая новорожденным птенцам (главным образом утятам) любой движущийся предмет соответствующих размеров (чучело, футбольный мяч, утку другого вида и т. д.), и при этом добивались запечатлевания птенцами этого предмета. В сочетании с рефлексом следования эти птенцы всюду следовали за запечатленным предметом. К. Лоренц произвел запечатлевание на себя выводка инкубаторных гусят, которые потом всюду следовали за ним и сопровождали его в воде при купании
Поскольку запечатлевание запоминается молодым животным весьма надолго", нередко на всю жизнь, при указанных выше экспериментах создавались поведенческие аномалии. Животные считали запечатленный предмет (или человека) не только своей матерью, но и сочленом популяции своего вида. В природе же, как правило, в момент рождения или вылупления около новорожденного оказывается мать или оба родителя. Именно их и запечатлевает молодое животное в первые часы и даже минуты своей жизни (так называемый чувствительный период). Таким образом, период «запечатлевания» представляется нам очень важным этапом — началом сигнальной преемственности у высших позвоночных. Дальше следует целая система воспитания (обучения) этих молодых животных, включающая подражание, следование, целый ряд сигналов, а нередко поощрения и наказания. У некоторых позвоночных животных этот период обучения длится недолго, а у других весьма длительное время.
У представителей класса рыб сигнальная преемственность, как правило, отсутствует, хотя, как было показано выше, при групповом обучении, обучение в стаях происходит здесь очень широко. Следует отметить, что у тех немногих видов рыб, для которых установлено семейное поведение, можно наблюдать элементы сигнальной преемственности, которые возникли в этом классе животных конвергентно под влиянием специфики взаимоотношений.
В классах амфибий и рептилий почти отсутствует активная забота о потомстве и соответственно нет семейных групп, не отмечена сигнальная преемственность. Можно лишь предполагать ее элементы у некоторых крокодилов.
У выводковых птиц птенцы с первого же дня жизни всюду следуют за матерью, подражая ей, копируя её движения и подчиняясь ее сигналам. Таким образом, они быстро научаются объектам и способам питания, а также распознаванию своих врагов и способам защиты (затаивания) при тревожных сигналах самки.
У птенцовых птиц из трех стадий ювенильной жизни, видимо можно составить два периода сигнальной преемственности. Первый — начальный период — от вылупления из яйца до покидания гнезда. Это период запечатлевания родителей и обстановки. Второй — активный период, когда птенцы оперены, выходят из гнезда, обучаются летать, и следуют за родителями, подчиняясь их сигналам. Именно в этот активный период, у птенцов образуется огромное количество условных рефлексов и формируются основные черты поведения взрослой птицы. При этом родители, конечно, бессознательно действуют часто по определенным программам.
По наблюдениям этологов, выводок поганок, покинув гнездо, чередует плавание и ныряние в воде с обогревом на спине родителей; это делается по программе, определяемой родителями. Птица сбрасывает с себя птенцов в воду и регулирует время их плавания, не давая им вернуться к себе на спину. По мере роста птенцов время их пребывания в воде птица увеличивает.
Самец большой синицы (Parus major) обучал своих летных птенцов маневрировать следующим образом:
он брал в кормушке кусок пищи и, подлетев к сидящим на ветке птенцам, садился около, а затем летел, маневрируя между ветвями; за ним летела вся стайка птенцов. Самец садился и давал кусок первому подлетевшему птенцу. Это повторялось многократно. Самка большого пестрого дятла, взяв из той же кормушки кусок хлеба, летела в сопровождении птенца к своей «кузнице», вставляла туда кусок и отлетала в сторону, как бы приучая птенца пользоваться «кузницей». Таких примеров можно привести большое количество. В случае опасности на птенцов действуют сигналы опасности родителей. Птенцы мухоловки-пеструшки не боялись ос и пчел в клетке и пытались их клевать. Если же жалящих насекомых впускали в клетку, где, кроме молодых, была и взрослая птица — кормилица, то все птицы, явно испуганные, сбивались в один угол, подальше от опасного насекомого. Очевидно, птенцы получали информацию об опасности от взрослой птицы.
 Многие черты в поведении птиц, которые входят в «видовой стереотип их
поведения» и считались ранее генетически закрепленными, после соответствующих
исследований оказываются приобретенными или формирующимися в онтогенезе на
базе опосредованного обучения и прежде всего «сигнальной преемственности». Это
было установлено у ряда видов птиц для особенностей песни и некоторых
акустических сигналов, имеющих в природе определенный видовой стереотип.
Многие черты в поведении птиц, которые входят в «видовой стереотип их
поведения» и считались ранее генетически закрепленными, после соответствующих
исследований оказываются приобретенными или формирующимися в онтогенезе на
базе опосредованного обучения и прежде всего «сигнальной преемственности». Это
было установлено у ряда видов птиц для особенностей песни и некоторых
акустических сигналов, имеющих в природе определенный видовой стереотип.
Именно опосредованным обучением можно объяснить относительно быстрое образование новых навыков у популяции в целом или у части популяции данного вида с резкими изменениями видовых стереотипов поведения. Можно привести несколько примеров.
Интересные изменения произошли за последние десятилетия в питании больших пестрых дятлов. Эти птицы, питавшиеся в основном древесными вредителями, частично перешли на выкармливание птенцов в теплые годы главным образом тлями, соскабливая их клювом с листьев и побегов. В холодные дождливые годы, когда прекращается размножение тлей, многие дятлы начинают хищничать, разоряя гнезда мелких птиц, преимущественно (но не только) дуплогнездников и, убивая их птенцов, питаются ими сами и кормят своих птенцов. Можно думать, что это явление так или иначе связано с уменьшением в культурных лесах старых деревьев, зараженных вредителями. Во всяком случае такое хищничество больших пестрых дятлов уже давно было известно в Западной Европе. На Украине оно было отмечено в начале 60-х годов и развивается все больше. Хищничество дятлов имеет место только в отношении птенцов семейства воробьиных. На взрослых птиц оно не распространяется. Во всяком случае, пришлось наблюдать, как самец-дятел перетаскал неоперенных птенцов из гнезда дрозда-белобровика, а самку дрозда, которая, защищая гнездо, налетала на дятла, он убил ударом клюва в голову, но есть ее не стал. По-видимому, имеет место «научение» дятлов от одного к другому и этой «науке» обучаются все более восточные дятлы». По наблюдениям, не все индивидуумы больших пестрых дятлов проявляют хищные наклонности. Было отмечено особенно большое хищничество дятла, но после отлова хищничающего самца разорение гнезд почти прекратилось, хотя в этом же районе держались и кормились еще две пары этих птиц. Не проявляла этих наклонностей и оставшаяся самка отловленного дятла. Весьма интересно, что хищное поведение дятлов отразилось на поведении других птиц: мухоловки серые и пестрые и дрозды - белобровики и рябинники начали нападать вблизи своих гнезд на больших пестрых дятлов и стараться прогнать их.
Американские этологи Фишер и Хинде описывают, как большие синицы в Англии научились протыкать, клювами картонные крышки у бутылок со сливками, которые торговцы оставляли утром у дверей домов своих клиентов, и выпивать часть сливок.Этот трюк, «изобретенный» отдельными птицами, переняли другие, так что он широко распространился в довольно большом районе, и молочники уже не осмеливались оставлять по утрам молоко у дверей домов. Вероятно, первые синицы научились этому методом проб и ошибок, а остальные — подражая первым». В поселках и городах нередко серьезной помехой хозяйкам бывают синицы-«форточницы», научившиеся расклевывать свертки с продуктами, вывешиваемые зимой за окно.
Следует вспомнить историю образования хищного поведения у новозеландского попугая кеа (Nestor notabilis). Эта некрупная птица (размером с ворону) живет в горных районах Новой Зеландии и питается обычно растительной пищей и мелкими животными. Однако в связи с развитием овцеводства некоторые кеа начали проявлять хищнические наклонности, нападая на овец и нанося им своим мощным клювом огромные раны в области спины. Овцы от этого погибают и их мясо поедается попугаями. Первые случаи такого хищничества были отмечены в 1868 г. Можно предполагать, что ему предшествовали случаи «чистки» овец попугаями от паразитов. При «чистке» ранки на коже овец кровоточили, и кровь пришлась, вероятно, по вкусу попугаям. Затем хищничество кеа приняло довольно широкие масштабы, причем, видимо, большое значение имело опосредованное обучение.
Интересный пример быстрого изменения особенностей поведения грача, гнездящегося в лесах Черноморского заповедника, всегда питались в основном насекомыми — вредителями сельского хозяйства и вдруг у обособленно живущей группы, гнездящейся на островке Соленоозерного участка, стали проявляться хищнические наклонности. Эта группа грачей жила в смешанной колонии с малой белой цаплей. У этих двух видов нередко возникали драки, во время которых разбивались яйца и расклевывались птенцы в гнездах. В 1964 г. грачи начали посещать о-в Орлов в Тендровском заливе и спокойно расклевывать лежащие в гнездах яйца чаек. Особенно большой урон они наносили гнездам черноголовых чаек, которые при тревоге не защищают своих кладок. Так «грачи-хищники», по выражению приспособились к добыче высококалорийного и легкодоступного корма.
В антропогенный период филогенеза происходят особенно быстрые изменения поведенческих адаптации животных. Человек так или иначе все сильнее вмешивается в жизнь животных нашей планеты. Он ускоряет естественный эволюционный процесс и придает ему новые направления. Одних животных он истребляет, другим изменяет условия жизни, третьих охраняет, одомашнивает и разводит. Очевидно, скорость возникновения морфологических адаптаций, если она не подкреплена искусственным отбором, не соответствует темпам развития деятельности человека, поэтому такие морфологические изменения очень редки. Основные адаптации у диких животных к этой деятельности человека развиваются главным образом по линиям общефизиологических (в первую очередь плодовитость) и поведенческих реакций. У животных ветви вторичноротых эти поведенческие адаптации создаются, прежде всего, в направлении опосредованного обучения (сигнальная преемственность и стайное обучение).
Многие преследуемые человеком промысловые животные выработали по отношению к нему сложные оборонительные особенности поведения. Они хорошо учитывают дальность действия современного охотничьего оружия, умело прячутся и запутывают свои следы. Нередко лисы, преследуемые гончими собаками в населенных местностях, используют шоссе и железные дороги для запутывания своих следов, что часто вызывает гибель собак. Волки и лисицы хорошо усвоили опасность капканов и запаха железа; в то же время лисы свободно роются на городских свалках и у туристских становищ среди консервных банок и других металлических предметов. При сокращении (от охотников) численности волков они делаются чрезвычайно осторожными, воспитывая это качество и в своем потомстве. Во многих местах стая волков обычно уходит от недоеденной добычи за десятки километров, что делает невозможным уничтожение их из засады, а также при помощи капканов и ядов.
Биологическое значение опосредованного обучения.
Биологическое значение опосредованного обучения во всем его комплексе очень велико. Оно проникает у птиц и млекопитающих даже в сексуальное и в материнское поведение. Например, как было показано в опытах с обезьянами, биологически нормальный самец, выращенный в изоляции от других особей своего вида, достигнув половой зрелости оказывается неспособным к нормальному для вида сексуальному поведению. Самка же, выращенная на искусственном кормлении под искусственной моделью «матери», не может правильно кормить своих детей и заботиться о них, а нередко уничтожает их.
Выходя из сказанного можно сделать вывод: кроме инстинкта здесь требуется еще кое – какое научение, осуществляемое в процессе общения с себе подобными.
Д. В. Радаков приводит интереснейшие наблюдения за поведенческими адаптациями животных к человеку. Так, он отмечает, что в 1968 г. в Актюбинской области(Россия) лисы выходили к железнодорожному полотну неподалеку от тех станций, на которых шла торговля копченой и вяленой рыбой, и ждали, когда из окон проходящего поезда пассажиры выбрасывали рыбные отходы. Он же описывает, как ласточки летают около идущего человека и следуют за повозкой или автомашиной, схватывая насекомых, вылетающих из травы. Подобно этому, сокол-чеглок ловит мелких птиц, выпугиваемых идущим поездом. Было отмечено, что ласточки летят за реактивным самолетом. едущим по дорожке, и ловят насекомых, поднятых из травы струёй газов двигателя.
 Следует
отметить, что относительно недавно появившаяся техника сначала вызывала резкую
оборонительную реакцию у многих животных, но затем в результате
опосредованного обучения довольно быстро превратилась не только в
индифферентный раздражитель, но в ряде случаев стала служить и положительным
пищевым условным раздражителем. Прежняя картина пахаря и сопровождающих его
грачей теперь заменилась трактором, идущим с плугами. Стрекотание такого
трактора привлекает животных, охотящихся за червями, личинками насекомых и
мелкими грызунами. За такими, тракторами движутся часто целые колонны грачей,
галок, ворон, речных и сизых чаек. Его сопровождают нередко и азартно охотящиеся
за грызунами лисицы. При этом, лисицы быстро устанавливают, в какой части поля
держится наиболее богатая почвенная фауна, и следуют за трактором именно на
этом участке.
Следует
отметить, что относительно недавно появившаяся техника сначала вызывала резкую
оборонительную реакцию у многих животных, но затем в результате
опосредованного обучения довольно быстро превратилась не только в
индифферентный раздражитель, но в ряде случаев стала служить и положительным
пищевым условным раздражителем. Прежняя картина пахаря и сопровождающих его
грачей теперь заменилась трактором, идущим с плугами. Стрекотание такого
трактора привлекает животных, охотящихся за червями, личинками насекомых и
мелкими грызунами. За такими, тракторами движутся часто целые колонны грачей,
галок, ворон, речных и сизых чаек. Его сопровождают нередко и азартно охотящиеся
за грызунами лисицы. При этом, лисицы быстро устанавливают, в какой части поля
держится наиболее богатая почвенная фауна, и следуют за трактором именно на
этом участке.
Интересные изменения происходят в поведении обыкновенного скворца (Sturnus vulgans). Эта птица, сильно тяготеющая к человеку, отличается необычайной пластичностью поведения. Гнезда скворца можно встретить и в дуплах деревьев, скворечниках, под застрехами домов в городах, их обнаруживали также в антенне действующего радиолокатора, в кузове работающею экскаватора и в других местах. Известно, что большие стаи скворцов остаются на зимовку и в Киеве. Д. В. Радаков описывает, как зимующие скворцы устраивают массовую ночевку на деревьях, растущих на центральной улице Киева. Тысячи скворцов облепляют здесь ветви деревьев, не опасаясь необычной обстановки этого большого густонаселенного города. Зимующие скворцы питаются в основном на городских свалках всевозможными отбросами и не упускают возможности поохотиться за мышевидными грызунами Обычно, пишет Радаков, в такой охоте принимают участие сразу несколько птиц, нанося грызуну удары клювом. Убив мышь, скворцы начинают ее расклевывать. Переход скворцов к такому хищничеству сказывается на их паразитофауне, поскольку на них все в большем количестве встречаются эктопаразиты мышевидных грызунов. Далее отмечает Радаков, что летом при выкармливании птенцов некоторые скворцы, живущие в населенных пунктах Киева и Киевской области, добывают пищу не только днем, но и в сумерках и даже ночью, охотясь у зажженных фонарей на слетающихся на свет насекомых.
Из всего вышесказанного можно сказать, что биологическое значение опосредованного обучения состоит в том, что особи популяций с опосредованным обучением быстрее приспосабливаются к изменчивым условиям окружающей среды.