Реферат: Приспособление организма к повышенной мышечной деятельности
Реферат
по биологии
на тему:
«Приспособление организма к повышенной мышечной деятельности»
2009
Введение
Нужно ли говорить, сколь разнообразен характер движения животных: и по биомеханической структуре, и по величине мышечных усилий, и по частоте циклов сокращения и расслабления, и по двигательному режиму. Нередко эволюционно близко стоящие друг к другу животные обладают совершенно различным характером движения. Достаточно сравнить скорость движения ерша или форели и ленивого налима или камбалы, ящерицы и черепахи, планирующий полет хищных птиц и гребной полет голубиных, воробьиных и куриных с частыми взмахами крыльев, двигательные возможности домашней и охотничьей собаки. Многие рыбы способны совершать миграции на очень большие расстояния. Например, угри, мигрирующие для нереста из Европы в Саргассово море через океан, совершают путь в 7000—8000 км, а лососевые из моря поднимаются по рекам против течения, нередко через пороги, и проходят 3000 — 4000 км со скоростью 4.2 км/ч. Огромные расстояния преодолевают и перелетные птицы: ласточки — до 1800 км, а буревестники — даже до 5000 км. Вместе с тем многие животные обладают не только большой выносливостью при движении, но и большой быстротой. Так, лиса может длительно* бежать со скоростью 35 км/ч, а гепард легко догоняет самую быструю антилопу, что недоступно собакам. Недаром в старину в Средней Азии и Индии гепардов использовали как охотничьих собак.
Большое разнообразие движений свойственно и человеку. Сравним работу мышц музыканта и тяжелоатлета-штангиста, быстрый бег спринтера и длительный марафонского бегуна, отточенные движения в гимнастике, фигурном катании или балете и труд землекопа. При этом на долю человека выпадают и очень большие мышечные нагрузки. Несмотря на развитие всех видов транспорта, механизацию, автоматизацию и роботизацию многих видов производства, человеку и в быту, и в трудовой деятельности приходится ходить на большие расстояния, быстро бегать на более короткие, поднимать и передвигать тяжести.
Естественно, что и человеку, и животному необходимо приспособиться к повышенной мышечной деятельности, чтобы она не вредила организму и результат ее был эффективен. Как и в рассмотренных выше реакциях, адаптация к повышенной мышечной деятельности может быть экстренной и кратковременной, длительной, приобретенной в результате систематической тренировки и видовой, закрепленной наследственностью.
Строение мышц и биохимия их работы
В связи с разнообразием движения мышцы животных и человека имеют разную форму и строение. Одни при работе могут проявлять большую силу при незначительном напряжении и долго не утомляться, другие отличаются быстротой сокращений, сильно напрягаются и быстро утомляются. Волокна, из которых построены мышцы, тоже неоднородны. Принято различать четыре типа их: медленные, быстрые гликолитические, быстрые оксидативно-гликолитические и промежуточные. При этом для волокон типа S характерны высокие возможности аэробных окислительных процессов и дыхательного генерирования АТФ, а для волокон типа F — большие потенциальные возможности мобилизации энергии АТФ и креатинкиназного и гликолитического ее ресинтеза. Третий и четвертый типы занимают промежуточное положение между первыми двумя.
В разные мышцы эти волокна входят в различных соотношениях. Одни мышцы, например предплечья, состоят преимущественно из волокон типа F; это быстро сокращающиеся, но скоро утомляющиеся мышцы. Другие, например камбаловидная мышца голени или мышцы, выполняющие более статическую функцию, как многие мышцы туловища, — в основном из волокон типа S; эти мышцы более медленно сокращаются, противостоят утомлению и способны к длительной работе умеренной интенсивности. Мышцы, содержащие поровну быстрые и медленные волокна или богатые волокнами FOG, обладают наиболее разнообразными функциональными возможностями: они способны и к быстрым сокращениям, и к длительной работе; это ряд мышц голени, бедра и плеча. Мышцы, в которых много волокон переходного типа I, наиболее пластичны, и при изменениях характера мышечной нагрузки возможно приспособительное превращение их в волокна одного из трех других типов. Кроме того, один тип волокон может приобрести некоторые черты другого при сохранении неизменными основных физиологических параметров, например волокон типов F и FOG.
Функциональные и биохимические особенности быстрых и медленных мышечных волокон
| Параметры | Тип F | Тип S |
| Возбудимость, MC | 0.2 | 10.0 |
| Функциональный ритм сокращений в 1 с | 30-50 | 10-15 |
| Скорость сокращения, мс | 84 | 95 |
| Скорость расслабления, мс | 167 | 205 |
| КФ, ммоль Р/кг | 12.6 | 11.0 |
| Гликоген, ммоль/кг | 28.0 | 24.4 |
| Активность, ммоль/: | ||
| АТФазы | 107.0 | 32.0 |
| креатинкипазы | 196.0 | 126.0 |
| фосфорилазы | 32.0 | 7.5 |
| малатдегидрогеназы | 240.0 | 355.0 |
| ферментов окисления жирных кислот | _ 1.1 | 4.2 |
| цитохромоксидазы, ммоль индофенолового синего/ | ■ 1.8 | 39.0 |
Мышечное сокращение, вызываемое двигательными нервными импульсами, является следствием взаимодействия сократительного белкового комплекса актомиозина, из которого построены фибриллы — тончайшие сократительные нити мышечного волокна, с АТФ. В результате того, что входящий в сократительный комплекс белок миозин обладает и ферментативными свойствами АТФазы, энергия, заключенная в фосфатных связях АТФ, без промежуточного образования тепла трансформируется в механическую энергию сокращения. Следовательно, мышечное сокращение сопровождается расходованием АТФ. Расслабление мышцы тоже требует затраты АТФ. В этом случае она идет на уборку ионов Ca, передающих информацию нервного импульса от двигательного нервного окончания к сократительному аппарату. А так как АТФ при мышечной деятельности все время потребляется, то она должна и восстанавливаться, ибо запасы ее в мышце невелики: их хватило бы всего на 2 — 3 с работы. По исчерпании этих запасов мышца не может более ни сокращаться, ни расслабляться и впадает в окоченение.
Ресинтез АТФ может происходить четырьмя путями. Самый первый по времени и наиболее быстрый — креатинкиназная реакция, т. е. перенесение богатой энергией фосфатной группы КФ на АДФ с образованием АТФ; но запасы КФ тоже невелики: их хватает лишь на обеспечение мышечной работы в течение 20—30 с. Второй путь — гликолиз; он дает возможность работать в течение минут и даже часа. Однако гликолиз имеет ряд недостатков: энергетически малоэффективен, приводит к наводнению организма молочной кислотой и сдвигу реакции внутренней среды в кислую сторону; кроме того, гликолизироваться может только глюкоза, а запасы гликогена в организме небезграничны. Достоинство же гликолиза в том, что он не требует от организма повышенного потребления кислорода. Третий путь ресинтеза АТФ — аэробное окисление различных веществ: углеводов, жирных кислот, глицерина, лишившихся своего азота остатков аминокислот, сопряженное с фосфорилированием АДФ. Положительное в нем — высокая энергетическая эффективность, конечные продукты его — безвредная для организма вода и легко удаляемая из него углекислота. Наконец, запасы источников энергии, способных подвергаться аэробному окислению, особенно жирных кислот, почти неиссякаемы. Но этот путь требует повышенного потребления кислорода. Четвертый путь — аварийный, невыгодный для организма и запускаемый только в крайних случаях. Это миокиназная реакция между двумя молекулами АДФ с образованием АТФ и АМФ: 2 АДФ АТФ + -f- АМФ. При этом АМФ теряет аминогруппу и превращается в инозиновую кислоту. Реакция эта необратима. Следовательно, тогда половина АДФ регенерируется в АТФ, а остальная АДФ представляет «издержки производства».
В условиях обычной, повседневной мышечной деятельности АТФ ресинтезируется путем аэробного окисления главным образом жирных кислот и в меньшей степени глюкозы. Но при интенсивной, повышенной мышечной деятельности это не всегда возможно или ограниченно. Для того чтобы пробежать 100 м со спортивной скоростью, надо затратить энергию, равную 135 кДж. Чтобы освободить такое количество ее путем аэробного окисления, организм должен потребить около 7 л кислорода. А человек, даже самый физически развитый, может поглотить Bt минуту 5 л кислорода или немного больше. Бег же на 100 м длится 10 — 12 с, причем многие спортсмены пробегают эту дистанцию на задержке дыхания, сделав глубокий вдох на старте. Таким образом, за время бега спортсмен получит не более 0.5—0.7 л кислорода. А нужно 7 л! Возникает кислородный дефицит, составляющий 90—95% от кислородного запроса, т. е. того количества кислорода, которое нужно организму в данных условиях. При других видах бега, более длительных, но менее интенсивных, кислородный дефицит меньше: при беге на 400—1500 м — 30—50%, при марафонском — 10, при беге на лыжах — 10—30%. Даже когда бежим к трамваю или автобусу или совершаем гигиенический «бег трусцой», у нас в организме образуется кислородный дефицит.
Приспособлением к такого рода мышечной деятельности служат креатинкиназная реакция и особенно гликолиз. Правда, гликолиз не обеспечивает полного восстановления всего израсходованного количества АТФ и КФ, -и содержание их в мышцах снижается: АТФ — на 15 — 35%, а КФ — в 2 раза больше. Однако и человек, и животное все же справляются с повышенной мышечной нагрузкой.
Восстановление нормальных, дорабочих биохимических соотношений, т. е. полный ресинтез АТФ, КФ и гликогена и устранение избытка молочной кислоты, происходит уже во время отдыха, когда организм «расплачивается» за анаэробное энергообеспечение мышечной деятельности. Эта «расплата», называемая кислородным долгом, выражается в повышенном поглощении кислорода в периоде отдыха, что делает возможным и окисление или превращение в гликоген молочной кислоты, и все репаративные синтезы. Кислородный долг всегда в той или иной мере больше кислородного дефицита. Повышенно поглощаемый кислород используется не только на энергообеспечение ресинтеза АТФ, КФ, гликогена и устранение избытка молочной кислоты, но и на полное восстановление биохимических соотношений в мышцах, нарушенных их повышенной деятельностью. Если во время мышечной работы кислородный запрос удовлетворяется не полностью, то миоглобин теряет свой кислород, повышенно разрушаются белки, фосфолипиды и даже некоторые субклеточные структуры, например часть митохондрий. Все это требует восстановления, а значит, дополнительного поглощения кислорода, являющегося как бы «процентами» за долг, которые тоже надо оплатить.
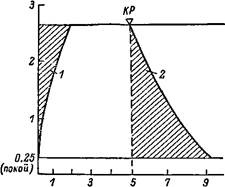
Соотношение кислородного дефицита и кислородного долга при интенсивной мышечной деятельности человека. По оси абсцисс — длительность работы, с; по оси ординат — потребление Ог, л/мин, KP — конец работы
Экстренное приспособление
В начале всякой мышечной деятельности, при переходе к ней от состояния покоя, энергообеспечение ее анаэробно, так как для того, чтобы обогащенная кислородом кровь дошла от легких до мышц, нужно 6—8 с. Следовательно, в начале работы запускается прежде всего креатинкиназный механизм ресинтеза АТФ, а затем — гликолиз. Чем работа интенсивнее, тем этот «пусковой период» значительнее и длительнее. При работе максимальной интенсивности вся работа выполняется в условиях «пускового анаэробиоза». При работах меньшей интенсивности потребление кислорода организмом увеличивается — и параллельно со снижением интенсивности гликолиза возрастает дыхательный ресинтез АТФ. Если интенсивность работы невелика, то потребление кислорода достигает величин, удовлетворяющих кислородный запрос, и энергообеспечение становится безраздельно аэробным.
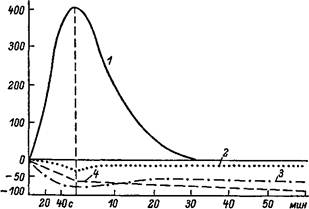
Расходование энергетических потенциалов мышцы в пусковой фазе и при работе в условиях устойчивого состояния. По оси абсцисс — время; по оси ординат — изменения от уровня покоя, Д%. 1 — молочная кислота, 2 — АТФ, 3 — КФ, 4 — гликоген
Потребление кислорода стабилизируется на определенной величине; интенсивность расходования мышечного гликогена снижается; основным источником энергии становятся жирные кислоты: молочная кислота частично окисляется, частично преобразуется в печени в гликоген; происходит ресинтез КФ; уровень КФ и АТФ стабилизируется несколько ниже исходного или даже достигает его. Это состояние энергообеспечения, называемое устойчивым, позволяет организму продолжать движение или работу весьма длительное время без заметных нарушений гомеостаза. Таким образом, экстренное приспособление к повышенной мышечной деятельности, осуществляемое только непосредственно при выполнении ее, сводится к обеспечению организма возможностью продолжать работу данной интенсивности во времени, к сведению до минимума ее стрессорного действия и сохранению необходимого для ее выполнения уровня гомеостаза.
Как же достигается это? Прежде всего обеспечением сокращающихся мышц источниками энергии. Как только к мышце поступает двигательный импульс и концентрация свободных ионов Ca в ней возрастает, происходит не только сокращение мышечного волокна, но и превращение малоактивной фосфорилазы b в высокоактивную форму а, что влечет за собой расщепление мышечного гликогена с образованием глюкозо-1-фосфата, подвергающегося затем гликолизу. Вызванное же мышечными сокращениями снижение концентрации АТФ направляет креатинкиназную реакцию вправо: АДФ + КФ АТФ + -f-крсатин. Таким образом, на какое-то время ресинтез АТФ обеспечен.
Но тут перед организмом встает новая важная задача: отбор мышечных волокон, которые при движении данной структуры, интенсивности и длительности должны сокращаться. Источником приходящих к мышце
двигательных импульсов являются специальные нервные клетки — мотонейроны спинного мозга. Их длинные отростки образуют двигательные нервы, причем каждый мотонейрон иннервирует от 5 — 10 до 100 мышечных волокон. Такой функциональный комплекс называется двигательной единицей. Все волокна, входящие в нее, под влиянием нервного импульса сокращаются одновременно с максимальной силой, подчиняясь закону «все или ничего». Поэтому сокращение мышцы в целом, сила его, зависит от количества возбуждаемых в данный момент двигательных единиц, что точно и тонко регулируется высшими отделами центральной нервной системы. Так как мышечные волокна разных типов иннервируются различными мотонейронами, нервная система избирательно рекрутирует те волокна, которые в данном случае нужны, и в тех соотношениях, которые обеспечат наиболее эффективное выполнение движения и продолжительность его во времени. Так, при работе малой интенсивности включаются главным образом и, в первую очередь, медленные волокна, обладающие большими возможностями аэробных окислительных процессов и дыхательного ресиитеза АТФ; при кратковременных нагрузках, по интенсивности близких к максимуму, — быстрые гликолитические.
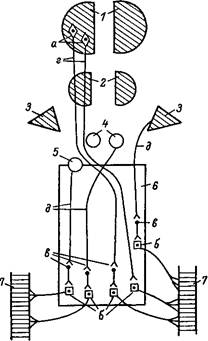
При длительных нагрузках в зависимости от их интенсивности к работающим медленным волокнам могут присоединяться в разное время и в разном количестве волокна быстрые и быстрые оксидативно-гликолитические. И все это в одной и той же мышце, содержащей волокна разных типов. Наконец, центральные влияния регулируют и число мышц, участвующих в данном движении, так как вовлечение в двигательную реакцию ненужных для нее мышц препятствует наиболее эффективному ее осуществлению и вызывает излишний расход энергии.
Возбуждение мотонейронов регулируется сложной системой нервных образований: корой головного мозга и таламусом через пирамидные пути, красными ядрами, ретикулярной формацией и мозжечком через экстрапирамидные пути. При этом между нейронами высших отделов нервной системы и мотонейронами стоят еще промежуточные вставочные нейроны, возбуждение или торможение которых в свою очередь отражается на функциональном состоянии мотонейронов.
Итак, запущено мышечное сокращение и отрегулирована структура движения — число участвующих в нем мышц, а в мышцах — различных типов волокон. Но движение должно продолжаться во времени, а для этого необходимы источники энергии. И в силу вступает влияние гормонов и симпатической нервной системы. Больше выделяется адреналина и норадреналина. Через систему цАМФ это приводит к расщеплению гликогена мышц и печени и повышенной доставке глюкозы к работающим мышцам. Несколько позднее тем же путем увеличиваются расщепление жиров и мобилизация жирных кислот. Поскольку возросшая мышечная деятельность является для организма стрессорной ситуацией, больше поступает в кровь и глюкокортикоидов, оказывающих пермиссивное действие на эффекты других гормонов. Наконец, рефлекторно усиливается активность систем дыхания и кровообращения, а значит, и доставка кислорода к работающим мышцам. Если мощность мышечной деятельности велика и сопровождается интенсивно идущим гликолизом, то вызываемое накоплением молочной кислоты смещение реакции внутренней среды в кислую сторону умеряется буферными системами крови и тканей.
Однако экстренное приспособление организма к мышечной деятельности хотя и позволяет продолжать работу во времени, но не предохраняет от быстрого наступления утомления, заставляющего организм прекратить работу или, во всяком случае, снизить ее интенсивность. Конечно, у организма есть возможности пустить в ход свои резервные силы при помощи центральной нервной системы. Например, высокий эмоциональный подъем в условиях увлекающего человека труда повышает работоспособность и отдаляет наступление утомления, как и поведение в экстремальных условиях. Стойкое же повышение работоспособности достигается долговременным приспособлением организма к различным видам повышенной мышечной деятельности.
Долговременное приспособление
Обратимся сначала к животным с разным характером движения и к разным мышцам одних и тех же животных, несущим неодинаковую нагрузку, т. е. к видовому, наследственно закрепленному приспособлению к повышенной мышечной деятельности. В мышцах разных беспозвоночных животных содержание АТФ и АДФ почти одинаково, тогда как количество аргининфосфата, заменяющего в них КФ, варьирует весьма широко, равным образом как и отношение фосфора аргининфосфата к фосфору АТФ и АДФ. А чем выше это отношение, тем больше возможности аргининкиназного ресинтеза АТФ; аргининкиназная реакция аналогична креатинкиназной. Содержание аргининфосфата и соотношение его с АТФ и АДФ выше у тех животных и в тех мышцах, которым свойственны быстрые движения. Так, мышцы брюшка речного рака выполняют более динамическую работу, чем мышцы клешни, хотя и быстро схватывающие, но длительно статически удерживающие добычу. Кузнечик больше прыгает, чем летает, а саранча, совершая большие перелеты, приземлившись, не прыгает, а спокойно передвигается на деревьях и хлебах, поедая их. Полевой скакун — очень быстрый жучок; навозный же жук ползает медленно. А насколько больше аргининфосфата и насколько выше отношение его содержания к содержанию АТФ и АДФ в мышцах конечностей кузнечика, чем у саранчи, или в грудных мышцах полевого скакуна по сравнению с теми же мышцами навозного жука. Вместе с тем активность окислительных ферментов в мышцах крыльев саранчи намного выше, чем у кузнечика, летающего недолго и недалеко.
Содержание богатых энергией фосфорных соединений, ммоль Р/кг, в мышцах беспозвоночных животных
| Животное | Мышцы | АТФ -f АДФ | Аргинин-фосфат | аргининфосфата Рдтф + лдф |
| Речной рак | Клешни | 12.2 | 13.7 | 1.13 |
| Брюшка | 13.6 | 16.0 | 1.18 | |
| Кузнечик | Конечностей | 13.6 | 37.4 | 2.75 |
| Крыльев | 13.0 | 24.3 | 1.89 | |
| Саранча | Конечностей | 10.8 | 21.0 | 1.94 |
| Крыльев | 13.0 | 20.0 | 1.54 | |
| Полевой скакун | Торакса | 11.3 | 39.0 | 3.45 |
| Навозный жук | » | 11.6 | 17.0 | 1.47 |
| Среднее | 12.5 | 23.5 | 1.90 |
В мышцах позвоночных животных концентрация АТФ тоже практически одинакова, тогда как содержание КФ и интенсивность гликолиза выше у более быстро движущихся животных: у лягушек выше, чем у жабы; у ужа выше, чем у черепахи; у быстрой мыши выше, чем у медлительной морской свинки, а у собаки и лисы, которые в быстроте бега могут соперничать друг с другом, эти показатели почти одинаковы. Широко варьируют и интенсивность дыхания мышечной ткани, и содержание миоглобина в мышцах. Первый параметр наиболее высок у лисы и собаки, второй — у кролика и лисы, способных к длительному бегу в большей степени, чем другие животные. Столь же велики интенсивность дыхания и содержание миоглобина в мышцах крота, способного к продолжительному напряженному рытью. К этому можно добавить, что у диких птиц содержание миоглобина выше, чем у домашних; у зайца выше, чем у кролика; у дикого козла выше, чем у домашнего; у охотничьих собак выше, чем у домашних.
Интересные данные обнаружены при сопоставлении возможностей гликолиза в мышцах разных животных и буферностью мышечной ткани: чем больше возможности образования молочной кислоты, тем выше буферность ткани. То же наблюдается и в отношении буферности крови. Эти особенности буферных свойств — приспособительные механизмы, направленные на поддержание гомеостаза при интенсивной мышечной деятельности, сопровождающейся значительным гликолизом.
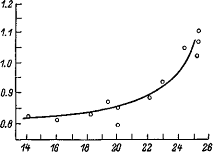
Соотношение между возможностями гликолиза в мышцах и их буферностью. По оси абсцисс — интенсивность гликолиза, мкмоль молочной кислоты на 1 г ткани в 1 мин; по оси ординат — мл 0.1 моль HCl, смещающие рН до 2.5
Активность миозиновой АТФазы, возможность мгновенно мобилизовать энергию АТФ и трансформировать ее в механическую энергию, у быстро и много движущейся форели и хищной трески выше, чем у озерных рыб — леща и налима. В ряду грызунов она убывает в соответствии с возможной быстротой движения. Активность креатинкиназы, содержание КФ и возможности гликолиза более значительны у весьма подвижной щуки, чем у менее подвижного подуста; у ящерицы выше, чем у черепахи; в летательных мышцах птиц с гребным полетом больше, чем у птиц с парящим полетом. Возможности дыхательного ресинтеза АТФ у нелетающих домашних птиц намного ниже, чем у диких. А среди последних величины этих параметров наиболее велики у перелетных птиц.
Если обратимся к разным мышцам одного и того же животного, то тоже обнаружим существенные различия в зависимости от характера работы, к которому приспособлена мышца. Мышца диафрагмы, всю жизнь ритмически сокращающаяся при дыхании и работающая в условиях устойчивого состояния обмена веществ, отличается наиболее низким содержанием КФ, гликогена и миозина, но в ней велики возможности дыхательного ресинтеза АТФ.
Биохимические особенности мышц некоторых позвоночных животных
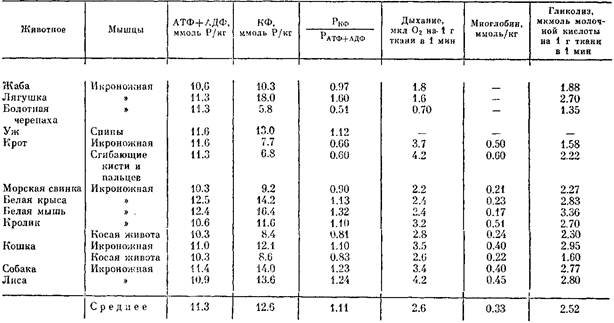
В мышцах конечностей, для которых характерны резкие переходы от покоя к сокращениям большой быстроты и силы, а нередко и большой продолжительности, содержание КФ, гликогена и миозина, а также активность АТФазы и гликолиза наиболее высоки, но велики возможности и дыхательного фосфорилирования. Наконец, косые мышцы живота, никогда с большой силой и быстротой не сокращающиеся, а несущие лишь небольшую нагрузку, содержат мало КФ и гликогена, АТФазная активность их низка. В процессах аэробного окисления они уступают диафрагме, а в гликолизе — мышцам конечностей. Активность миозиновой АТФазы и содержание сократительных белков в мышцах передних конечностей крота, совершающих большую и интенсивную работу при отрывании нор, значительно выше, чем в мышцах задних конечностей, служащих лишь для довольно медленного передвижения. Следовательно, высокая АТФазная активность нужна не только для быстрого сокращения мышц, но и для развития силовых напряжений.
Важным свойством мышц, отличающихся большой силой, является и их масса. У того же крота масса мышц предплечья намного больше, чем мышц голени. Вспомним, как мощны мышцы плечевого пояса у бизона или зубра. Сравним мышцы лошадей-тяжеловозов и предназначенных лишь для верховой езды или собственную мускулатуру с мышцами тяжелоатлета-штангиста.
В разных мышцах одного и того же животного неодинаково и содержание миоглобина: наиболее богаты им те мышцы, которые несут большую нагрузку при движении. Так, в мышцах конечностей оно выше, чем в мышцах брюшной стенки и спины. У птиц оно более значительно в больших грудных мышцах, чем в малых с меньшей нагрузкой при полете. У крота оно особенно велико в мышцах предплечья.
Рассмотренные данные позволяют выявить общую закономерность: чем большую и более длительную нагрузку несет мышца, чем выносливее она к длительной работе, тем выше в ней возможности дыхательного ресинтеза АТФ и тем лучше условия для обеспечения его. Для тех мышц, которым свойствен резкий переход от покоя к весьма интенсивной работе, выполняемой кратковременно, но близкой к максимуму мощности, характерны высокая АТФазная активность, значительное содержание КФ и большие возможности гликолиза. Наконец, для мышц, развивающих большие силовые напряжения, тоже свойственны большая
АТФазная активность, высокое содержание миозина и миоглобина, а также значительная общая масса мышцы.
Все описанные приспособления видовые; они произошли в процессе эволюции, закреплены наследственностью и отбором. Проявляются ли эти закономерности при индивидуальной длительной адаптации к повышенной мышечной деятельности?
Излюбленным объектом биохимиков при исследовании длительного приспособления к мышечной деятельности являются белые крысы. Они могут выполнять самые различные мышечные нагрузки: бег в колесе или на механически движущейся дорожке с различной скоростью и длительностью; продолжительное многочасовое плавание; более короткое, но более интенсивное плавание с грузом, подвешенным под брюхо; силовые нагрузки: свободно висеть на вертикальном стержне или с «рюкзачком», наполненным дробью, массу которого можно изменять. Тренировка животных строится по тем же принципам, что и спортсменов: строго регламентируются величина нагрузки и время последующего отдыха, возрастание нагрузки на протяжении тренировочного процесса и т. д., а вся тренировка длится от 1 до 4 мес.
Закономерность, выявленную при рассмотрении видового приспособления к повышенной мышечной деятельности, полностью подтверждают данные, полученные в результате тренировки разными нагрузками. Прежде всего обращает на себя внимание, что вызываемые разной тренировкой биохимические изменения в мышцах имеют неодинаковое количественное выражение. Долговременное приспособление к длительной мышечной деятельности умеренной мощности сопровождается наиболее значительным возрастанием возможностей дыхательного генерирования АТФ; приспособление к кратковременной мышечной деятельности, по мощности близкой к максимуму, — наибольшим увеличением возможностей анаэробного генерирования АТФ, а при приспособлении к силовым напряжениям основные изменения касаются различных мышечных белков. Наиболее значительно возрастают возможности мобилизации АТФ, сокращения и расслабления мышцы, масса мышц и их волокон. С помощью микробиопсии, т. е. извлечения специальной иглой небольших проб мышцы, что совершенно безболезненно, та же направленность биохимических изменений установлена и у тренирующихся спортсменов разного профиля.
Таким образом,- приспособление организма к повышенной мышечной работе характеризуется увеличением энергетического потенциала мышц и возможностей использования его при мышечной деятельности. При всех видах тренировки отсутствуют изменения содержания АТФ в мышцах. Объясняется это тем, что благодаря более энергичному расщеплению и ресинтезу АТФ при сокращении мышц «оборачиваемость» ее в тренированном организме возрастает и в этих условиях того же количества АТФ хватает на энергообеспечение значительно большей работы.
Так как при долговременном приспособлении к мышечной деятельности увеличивается содержание многих мышечных белков, можем констатировать повышение синтеза их, обусловленное активацией генома и всего белоксинтезирующего аппарата. При этом не только усиливается синтез белков, но и происходит дополнительная сборка субклеточных образований, в частности митохондрий, число которых в мышце возрастает. Такая сборка ведется по новому плану, задаваемому характером мышечной деятельности. Об этом можем судить по изменению соотношения компонентов дыхательной цепи митохондрий, передающей электроны и протоны от окисляемых веществ на кислород.
В результате этого возрастают возможности окисления различных субстратов, но для разных веществ неодинаково: под влиянием тренировки длительными нагрузками окисление ПВК увеличивается на 200%, янтарной кислоты — на 40, глицерофосфата — на 28%. При тренировке скоростными нагрузками — соответственно на 100, 50 и 3%.
Как же происходят все эти приспособительные изменения, каков их молекулярный механизм? И почему эти изменения так удивительно совпадают с интересами организма, с тем, что необходимо ему при изменившемся в том или ином направлении характере мышечной деятельности? Мы уже знаем правило В. А. Энгельгардта о том, что всякая реакция расщепления вызывает или усиливает в организме реакцию, производящую ресинтез. Во время мышечной деятельности расщепляются прежде всего источники энергии: АТФ, КФ, гликоген. Значит, при переходе от работы к покою должен осуществляться ресинтез их; причем, чем более интенсивными и значительными были затраты, тем энергичнее должен быть и ресинтез.
Еще в начале 50-х гг. было установлено, что расходуемые мышцей вещества восстанавливаются не до исходного уровня, а через известное время даже превышают его. Вслед за восстановлением идет фаза сверхвосстановления, а затем постепенное волнообразное возвращение к первоначальному уровню. Фаза сверхвосстановления не очень длительна: от нескольких часов до 1—2 сут. Но если каждую повторную работу начинать в этой фазе, то уровень источников энергии будет все время возрастать и в конце концов станет стабильно повышенным.
Размеры сверхвосстановления зависят от величины и интенсивности расходования. При скоростных нагрузках весьма интенсивно расходуется КФ, поэтому уровень его при тренировке этими нагрузками в наибольшей степени возрастает. При длительных и скоростных нагрузках интенсивно тратится мышечный гликоген, а при силовых нагрузках его требуется немного. Поэтому в первых двух случаях тренировка приводит к значительно большему повышению содержания гликогена, чем тренировка силовыми нагрузками. Вместе с тем при силовых нагрузках мышечных белков расходуется существенно больше, чем при скоростных и тем более длительных. Поэтому при тренировке силы и наблюдается наибольший синтез белков.
Правилу В. А. Энгельгардта подчиняются все вещества, используемые или разрушаемые при мышечной деятельности, кроме АТФ. Дело в том, что АТФ все время расходуется и ресинтезируется. Правда, если работа была очень интенсивной, уровень АТФ в мышцах снижается. В этом случае сразу по окончании работы может наблюдаться очень короткая и слабо выраженная фаза сверхвосстановления, но она быстро проходит, так как для всех репарационных синтезов в период отдыха необходима затрата АТФ. В это время продолжаются и расходование АТФ, и ее энергичный ресинтез, которые уравновешивают друг друга и стабилизируют содержание АТФ на уровне, свойственном состоянию покоя.
Механизм приспособительных изменений мышц к повышенной их деятельности не ограничивается явлением сверхвосстановления. В мышце возрастает содержание ряда веществ, в первую очередь многих структурных и ферментных белков, которые не расходуются или используются очень мало. В этом случае их синтез стимулируется дерепрессией генов, где закодирована структура этих белков. Предполагается, что дерепрессорами и индукторами синтеза могут быть различные вещества, прежде всего нормальные продукты обмена веществ, а также некоторые гормоны: глюкокортикоиды, инсулин и др. Для синтеза фррментных белков эту роль, видимо, может играть повышение концентрации субстратов катализируемых ими реакций. В пользу этого говорит первоочередное и более значительное увеличение синтеза тех ферментов, которые обладают меньшей «пропускной способностью» и катализируют реакции, наиболее существенные при данном роде мышечной деятельности.
Так как синтез белков требует значительной затраты энергии, начинается он не сразу, а по достижении сверхвосстановления содержания источников энергии. Сама мышечная деятельность и вызываемое ею нарушение гомеостаза создают предпосылки для повышения в процессе тренировки содержания источников энергии, улучшения условий для усиления синтеза белков.
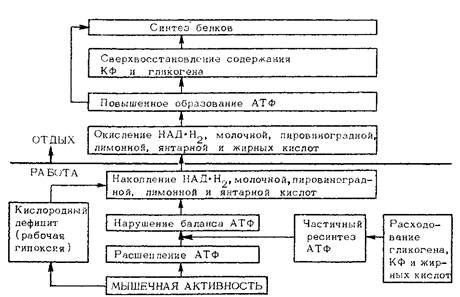
Последовательность репарационных биосинтезов в мышцах во время отдыха после работы.
Насколько же стойки все эти молекулярные приспособительные изменения? Оказывается, стойкость их имеет предел, при этом разные качества мышечной деятельности снижаются через разное время. Для поддержания их на должной высоте необходима активная мышечная деятельность. Об этом свидетельствуют и наблюдения на людях, и опыты с тренировкой животных. Ведь хорошо известно, что спортсмен, переставший тренироваться, теряет свою «спортивную форму», его организм становится менее приспособленным к выполнению тех больших физических нагрузок, которые доступны были ему ранее. В первую очередь уменьшается возможность проявления максимальной быстроты, затем — силы при динамической работе, позднее — силы при статических мышечных напряжениях, а выносливость к длительным нагрузкам умеренной интенсивности сохраняется наиболее долго. То же показывают и результаты опытов на животных, только сроки у них намного короче: если у человека — месяцы, то у лабораторных крыс — несколько суток. При этом, чем более длительной была тренировка, тем дольше сохраняются достигнутые адаптационные изменения. Но они не становятся наследственными, во всяком случае на протяжении ближайших поколений.
А как же быть с видовыми приспособлениями к повышенной мышечной деятельности различного характера? Обратимся к предположениям и теоретическим построениям, поскольку экспериментальных данных здесь немного. Так, в 1939 г. американский биохимик В. Блур провел опыты с тренировкой нескольких поколений крыс, исследуя у них содержание фосфолштидов в мышцах, которое под влиянием тренировки повышается. Каждое новое поколение рождалось с обычным количеством этих веществ в мышцах и сохраняло такой уровень до тех пор, пока не начиналась тренировка. Но уже с 3—4-го поколения содержание фосфолипидов при тренировке стало повышаться раньше, чем у их «отцов, дедов и прадедов». Иначе говоря, через несколько поколений возросла скорость приспособления к повышенной мышечной деятельности, хотя сама адаптация не стала наследственно закрепленной. Это уже шаг к видовому приспособлению.
Как и что наследственно закрепляется при видовом приспособлении, еще сказать нельзя. Ясно одно: дело не в мутации генов, кодирующих структуру различных синтезируемых белков. Ведь ни при индивидуальном долговременном приспособлении к повышенной мышечной деятельности, ни в видовых различиях химизма мышц животных с разным характером движения нет, а значит, и не синтезируется никаких новых для организма белков. Все белки, о которых шла речь, закодированы в геноме и у разных животных не имеют принципиальных отличий. Но вот объем их синтеза может быть существенно различным. А из этого следует, что причины видовых приспособительных особенностей мышц надо искать в наследственных, закрепленных отбором механизмах регуляции функциональной активности генома. Но это еще дело не сегодняшнего, а, так сказать, завтрашнего дня.
Конечно, приспособление организма к повышенной мышечной деятельности не ограничивается рассмотренными молекулярными изменениями мышц. Оно касается и центральной нервной системы, и гормональной регуляции физиологических функций, систем дыхания, кровообращения и других сторон жизнедеятельности организма.
Приспособление к ограничению подвижности
Весьма неблагоприятное влияние на организм оказывает ограничение подвижности, связанное с условиями труда, длительным постельным режимом при заболеваниях и травмах, а тем более с космическими полетами, где присоединяется еще влияние невесомости. Уменьшение работы мышц резко ослабляет афферентную импульсацию и приводит к угнетению возбудимости центральной нервной системы, реактивность которой вообще снижается. Падает тонус кровеносных сосудов, ослабляется работа сердца, атрофируются мышцы, меньше становится в них сократительного белкового комплекса — актомиозина. Угнетается синтез белков мышц и внутренних органов, а расщепление их усиливается, в результате чего организм в повышенном количестве теряет азот, серу и фосфор, а вследствие частичной деминерализации костей — и кальций. Уменьшение синтеза белков влечет за собой снижение приспособляемости организма к повреждающим факторам и падение устойчивости его к гипоксии, проникающей радиации, охлаждению и перегреванию.
Естественно, что при разных причинах и формах гипокинезии все эти изменения выражены в различной степени. Наиболее значительны они при многомесячном строгом постельном режиме и особенно при космических полетах.
Единственным средством «приспособиться» к длительному ограничению подвижности и противостоять его вредоносному влиянию является активная мышечная деятельность. Если гипокинезия связана с условиями труда, следует больше внимания уделять утренней и производственной гимнастике, предпочитать транспорту хождение пешком, заняться каким-либо видом спорта или посещать группы здоровья. При вынужденном постельном режиме необходима лечебная физкультура, а при космических полетах, как известно, члены экипажа систематически выполняют значительные физические нагрузки, комплексы специальных физических упражнений. Соблюдение всего этого спасает организм от отрицательного влияния гипокинезии, сохраняет здоровье и высокую работоспособность.
| Физиология бега | |
|
Содержание 1 Бег и его место в классификации физических упражнений 2 Спортивный результат в беге и физическое развитие 3 состояние ЦНС и сенсорных ... Эта система типична для кратковременных усилий, например, для первой половины бега на дистанцию 100 м. Если мы стартуем внезапно из состояния покоя, наши мышцы начинают расходовать ... В работающих мышцах бегуна при утомлении происходит исчерпание запасов энергетических субстратов (АТФ, КФ, гликоген), накапливаются продукты распада (молочная кислота, кетоновые ... |
Раздел: Рефераты по физкультуре и спорту Тип: контрольная работа |
| Бодибилдинг | |
|
Индивидуальное задание по теме Выполнил тудент II курса факультета ПМиКТ Руммо Евгений Андреевич Вологодский государственный педагогический ... Длина мышцы зависит от длины мышечных волокон. Неизрасходованное энергетическое "топливо" будет откладываться в виде жировых отложений, в виде гликогена в печени и в мышцах, а также в мышечных клетках в виде дополнительного ... |
Раздел: Рефераты по физкультуре и спорту Тип: реферат |
| Мышечная система человека | |
|
Реферат "Мышечная система" Введение Мышцы (musculi) - активная часть двигательного аппарата человека. Кости, связки, фасции образуют его пассивную ... Сократимая часть мышцы, образованная мышечными волокнами, с обоих концов переходит в сухожилие. При физической тренировке происходит утолщение мышечных волокон и увеличиваются их энергетические ресурсы. |
Раздел: Рефераты по биологии Тип: реферат |
| Психологическая подготовка к рукопашному бою | |
|
Психологическая подготовка к рукопашному бою Алексей Алексеевич Кадочников Аннотация В книге представлены основные методы психологической подготовки ... Для тренеров диагноз ясен: надо срочно снизить нагрузку в двух-трех тренировках. Предложенные приемы мышечной релаксации можно проделать без длительной тренировки, эффект их неоспорим. |
Раздел: Рефераты по физкультуре и спорту Тип: книга |
| Параметры функционирования митоКАТФ у животных с различной ... | |
|
Содержание Список сокращение Введение Глава 1. Обзор литературы 1.1 Системы транспорта калия в митохондриях 1.1.1 Транспорт калия в митохондрии 1.1.2 ... 1) механизм, при котором усиление входа К+ в МХ ведет к набуханию матрикса, что предупреждает распад АТФ; 2) механизм, основанный на снижении концентрации Са2+ в МХ, что уменьшает ... Для этого выделенный из МХ печени крысы и очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа наносили на ДДС-ПААГ электрофорез с 10%-ым ПААГ гелем с нагрузкой на ... |
Раздел: Рефераты по медицине Тип: дипломная работа |