Дипломная работа: Проблема сохранения биологического разнообразия
Дипломная работа
«Проблема сохранения биологического разнообразия»
Введение
Актуальность исследования. Современные тенденции развития географической науки требуют разработки научных основ инвентаризации и оценки природных, в частности биологических, ресурсов территории. До последнего времени при изучении ландшафта, анализе его структуры, функционирования и условий обособления основное внимание уделялось абиотическим компонентам. Животное население в силу своей подвижности и пластичности, а, следовательно, и сложности изучения либо не рассматривалось вообще, либо механически проецировалось на растительные сообщества. Исходя из этого, следует отметить весомую актуальность исследования животного компонента ландшафтов как наиболее мобильной системы, отражающей любые изменения как внешней, так и внутренней среды природного комплекса.
Особое значение такие исследования приобретают в условиях высокогорных ландшафтов, в частности, Западного Кавказа, которые при современном уровне антропогенной трансформации природных комплексов, играют важную роль в сохранении генофонда биологического разнообразия и выступают в качестве эталонных участков биосферы (Второв, 1968).
Проблема сохранения биологического разнообразия является одним из приоритетных направлений в современной биогеографии. В настоящее время возрастает антропогенное воздействие на биотические компоненты природы, создается угроза разрушения биоэкосистем и биологического разнообразия биосферы. Эта проблема актуальна в региональных исследованиях горных территорий, хрупких и восприимчивых к человеческой деятельности, для которых характерны быстрое сужение качественной среды обитания, утрата биоразнообразия.
Приходится признать, что большинство биогеографических исследований в пределах Западного Кавказа (северный склон) посвящены в основном изучению особенностей распространения биоты в пределах высотных поясов (количественная и качественная характеристики, оценка состояния растительного покрова и т.д.). Горизонтальная дифференциация растительного покрова и хортобионтов, с учетом среды их обитания в пределах морфологических единиц высокогорных ландшафтов изучена слабо. В связи с этим возрастает актуальность изучения организации высокогорных сообществ с помощью ландшафтного подхода, сущностью которого является анализ взаимосвязей всех элементов и компонентов природы территории. А в зависимости от набора морфологических единиц “географического экотона” (местностей и урочищ) и соотношения по площади их лесных и луговых сообществ можно определять границу среднегорных лесных и высокогорных луговых ландшафтов, относя переходный пояс к тем или иным горным ландшафтам. Также не менее актуальными и слабо изученными являются средоформирущие факторы и среда биоты, в частности, группы беспозвоночных. Использование индикационных геоботанических методов позволяет анализировать состояние травянистой растительности, привязанных к ней хортобионтов, а также выявлять степень антропогенной трансформации высокогорий.
Целью работы явилось изучение особенностей внутриландшпафтной дифференциации беспозвоночных в пределах горного экотона лесных и луговых высокогорий как индикатора границ этих групп ландшафтов, выявление их региональных различий.
В ходе работы были поставлены следующие задачи:
1) создание ландшафтных карт ключевых участков ландшафтов исследования;
2) выявление и изучение особенностей внутриландшафтной дифференциации беспозвоночных значимых доминантных групп с учетом особенностей среды обитания;
3) выявление различий групп беспозвоночных горного экотона в разных регионах Западного Кавказа;
4) использование данных по беспозвоночным в качестве биоиндикации антропогенной нарушенности.
Объектом исследования явились насекомые урочищ ландшафтов различных видов экотона высокогорий Западного Кавказа, в частности Orthoptera, Hemiptera и Homoptera и Coleoptera.
Предметом изучения стала внутриландшафтная дифференциация беспозвоночных с учетом среды их обитания в разных видах горного экотона.
Материалы и методы исследования. Работа проводилась в рамках развиваемого в СГУ научного направления «Биотика ландшафта». В основу работы положены материалы собственных полевых и камеральных исследований 2003-2006 годов.
Научная новизна исследования:
- исследованы особенности горизонтальной (по морфологическим единицам ландшафта) и вертикальной (по высотным поясам) дифференциации беспозвоночных высокогорных ландшафтов в зоне горного экотона;
- определены тенденции изменения структуры беспозвоночных в зависимости от морфологии ландшафтов и среды их обитания;
- получены первичные критерии изменения структуры населения беспозвоночных в зависимости от степени антропогенной нарушенности территории.
Методология и теория исследования базируется на ландшафтном и ландшафтно-экологическом подходах, учении о горном ландшафте: его морфологии (геоботанический пояс, местность, урочище, фация), биоразнообразии, среде жизни беспозвоночных, средоформирующих факторах, особенностях границ ландшафтных поясов и контурных границ морфологических единиц ландшафтов.
Методика исследования основана на ландшафтном подходе с использованием группы методов: математических, метеорологических, ландшафтного и функционального анализа, а также картографических.
Апробация работы и публикации. Основные положения работы были представлены на: 50-й, 51-ой и 52-ой научно-методических конференциях “Университетская наука - региону” (апрель 2005, 2006, 2007) Ставрополь; Международной научно-практической конференции “Проблемы экологической безопасности и сохранения природно-ресурсного потенциала” (29-30.09.2006) Ставрополь.
Объем и структура работы. Магистерская диссертация состоит из введения, трех глав, заключения, списка литературы. Она содержит 86 страниц текста, включает 29 рисунков, 4 таблицы. Список литературы включает 93 источника.
Глава I.
История вопроса и теоретические положения ландшафтно-экологического подхода
Общеметодологическим подходом исследования является ландшафтный подход, который все еще не получил должного развития, особенно в части практической реализации. О его сущности и важности неоднократно писали классики физической географии XX века: Д.Л. Арманд, Н.А. Гвоздецкий, И.П. Герасимов, А.Г. Исаченко, К.К. Марков, Ф.Н. Мильков, В.С. Преображенский, Н.А. Солнцев, В.Б. Сочава и др.
Сущность ландшафтного подхода заключается, во-первых, в учете индивидуальности природы земной поверхности, организованной в сочетания ПТК, образующих относительно однородные по генезису территории, называемые ландшафтами; во-вторых, в учете их пространственно-временной иерархической структуры; в-третьих, учете причинно-следственных взаимосвязей между отдельными компонентами. Отсюда важное следствие: что ландшафтный подход может выступать в качестве общеметодологической базы или составной части в обосновании и реализации социальной, экономической, экологической, военной и других “политик” и программ. Без него (ландшафтного подхода) самые совершенные программы будут ущербными (Дьяконов, 2005).
В настоящем исследовании он включает учение о горном ландшафте и понятии «горного экотона», а также понятие ландшафтной среды, являющейся составной частью географической среды.
1.1. Учение о горном ландшафте
Теоретической основой настоящего исследования является учение о горном ландшафте.
Горным природным ландшафтом называется генетически однородный участок ландшафтного пояса, расположенного в пределах определенного высотного яруса гор, имеющего одинаковый геологический фундамент, один тип рельефа, одинаковый климат, определенный набор высотных геоботанических поясов и других морфологических единиц (Шальнев, 2004).
Понятие о горном ландшафте, сложившееся в географической литературе, не однозначно. Это связано с тем, что зональные закономерности претерпевают здесь значительные изменения в связи с ярусностью гор. Ярусность отражает этапы формирования горной системы, возраст отдельных ее частей, характер экзогенного расчленения, климатическую дифференциацию. В пределах Западного Кавказа выделяется три ландшафтных яруса: предгорий и низкогорий, среднегорий и высокогорий. Для каждого высотного яруса характерен свой набор или спектр ландшафтных высотных поясов.
Исследованием отдельных территорий данного региона занимались Н.А. Гвоздецкий, А.Г.Исаченко, А.Е.Федина и др. Есть публикации фрагментов ландшафтных карт (Исаченко, 1965; Шальнев, 1971, 1973; Шальнев, Кондратьева, 1994 и др.). Ландшафтная карта Кавказа, созданная Н.Л. Беручашвили (1979), включала и территорию Северного Кавказа. Этот же автор в 1995 г. публикует книгу о ландшафтах Кавказа в электронном варианте. Ландшафты Восточного Кавказа изучал В.В. Братков (1992). Обзорную ландшафтную карту Северного Кавказа создают географы Ростовского университета (Чупахин, 1974). Есть публикации по изучению ландшафтов Чечено-Ингушетии (Волынкин, Доценко, 1977, 1979) и Предгорного Дагестана (Атаев, 1988). Опубликована ландшафтная карта в Атласе Дагестана (1999).
Высотно-поясное деление является «яблоком раздора» в морфологии горных ландшафтов. Одни авторы (Гвоздецкий, 1963, Чупахин, 1974) считают высотный пояс (зону) подразделением высшего ранга, созвучным с понятием подзона на равнине. Другие отождествляют его с понятием ландшафта (Михайлов, 1962) или микроландшафта (Тумаджанов, 1963). Многие географы (Геренчук и др., 1963; Исаченко, 1991) относят высотный пояс к категории морфологических единиц, выделяя его наряду с фациями, урочищами и местностями. Есть и другие терминологические различия. В одном случае, употребляя термин «высотный пояс», имеют в виду пояс растительности (Исаченко, 1991). У других авторов используется другой термин – ландшафтный пояс. Местности образуют «своего рода пояса, которые следует называть ландшафтными поясами, а не поясами растительности» (Геренчук и др., 1963, с.38).
На взгляд В.А. Шальнева, это разные понятия и они имеют право на существование в учении о горных ландшафтах. Так, для геоботаников высотный пояс растительности является устоявшимся понятием и под ним «подразумевается более или менее широкая горизонтальная полоса растительности в горах с господством или одного типа растительности или нескольких, закономерно чередующихся» (Лавренко, 1964, с.190). Такие высотные пояса формируются в пределах определенных высотных уровней. Например, пояс хвойных лесов формируется в диапазоне абсолютных высот 1300 – 2100 м над у.м., субальпийских лугов – 2200-2600м. Границы растительных поясов под влиянием изменяющихся с высотой различных факторов (главным образом климатических) фиксируют качественные изменения одного или нескольких компонентов среды – растительности, почв, климата (Щукина, 1960; Шальнев, Кондратьева, 1994). Такие высотные пояса лучше называть геоботаническими, так как их внешний «портрет» определяет растительность. Высотный геоботанический пояс – пояс, относящийся к категории внутриландшафтной вертикальной дифференциации и формирующийся под влиянием изменения с высотой климатических факторов и фиксирующий качественные изменения одного (растительности) или нескольких компонентов среды – растительности, почв, климата (Щукина, 1960).
Геоботанические высотные пояса неоднородны по своей сущности. Выделяются доминантные и переходные пояса. Доминантный пояс относится к числу основных высотных поясов, создающих специфический пейзаж того или иного участка гор. Он формируется в условиях господствующих климатогенных условий (климатогенное поле однородности) и имеет значительное вертикальное развитие и горизонтальную протяженность. Переходный высотный пояс формируется в результате количественных и качественных изменений единиц климатогенных полей при переходе от одного доминантного пояса к другому (Шальнев, Кондратьева, 1994). Поэтому для него характерно преобладание не одного, а нескольких типов растительности и других компонентов природы, формирующихся на склонах разной экспозиции (сосновых лесов и нагорных ксерофитов или сосновых лесов, березовых криволесий и субальпийских лугов). Переходные пояса сочетают в себе свойства двух пограничных доминантных поясов и отражают закономерности, получившие название горного экотона (Шальнев, Юрин, 1997).
Ландшафтный пояс – более объемное понятие, чем геоботанический. Он формируется в пределах округов или провинций и приурочен к конкретному ландшафтному ярусу гор. В связи с этим имеет определенную однородность в геоморфологическом и литогенном отношениях. Климатогенные и биогенные компоненты более разнородны и представлены набором генетически однородных геоботанических поясов – доминантного (он дает название ландшафтному поясу) и одного–двух переходных. Ландшафтные пояса, по мнению В.А. Шальнева, являются региональными природными образованиями. С ними связано распространение определенных видов ландшафтов. Геоботанические же пояса относятся к категории внутриландшафтных подразделений вертикальной дифференциации. С учетом названных подходов была составлена карта ландшафтов Карачаево-Черкесской республики.
Местность – сложно устроенная морфологическая часть ландшафта, которая в одном и том же ландшафте обычно несколько отличается геолого-геоморфологическим строением и, вследствие, этого набором урочищ, при этом нередко фоновые урочища остаются теми же, а изменения касаются субдоминантных или же дополняющих урочищ (Хрусталев, 2000).
Урочище – природный территориальный комплекс, состоящий из систем и их групп, динамически и территориально связанных фаций; обычно они формируются на основе какой-либо одной мезоформы рельефа (Хрусталев, 2000).
Фация – самая мелкая таксономическая единица природно-территориального комплекса, формирующаяся на основе микроклиматических показателей и растительного покрова (Хрусталев, 2000).
1.2. Понятие “горного экотона”
В современной географии одним из перспективных и интересных направлений стало изучение экотонов. Само понятие «экотона» как переходного пространства между фитоценозами, обладающего специфическими свойствами (повышенным биоразнообразием, обилием организмов) было предложено Ф. Клементсом в 1928. Позже проблема границ в природе и переходных «буферных» пространств привлекла многих других ученных, в том числе и географов. Среди них В.Б. Сочава, Д.Л. Арманд, В.С. Залетаев, Э.Г. Коломыц, В.А. Николаев и др. Из географов на проблемы экотона впервые обратил внимание В.Б. Сочава [1978]. Он применил понятие «экотон» для обозначения буферного сообщества, а позже как «переходную полосу между двумя регионами, или двумя выделами геомеров». Ф.Н. Мильков [1986] дал более развернутое определение. Под экотоном он понимает переходную полосу между смежными ландшафтными комплексами, для которой характерна повышенная интенсивность обмена веществом и энергией, разнообразие экологических условий и, как следствие, высокая концентрация органической жизни.
В тоже время, проблемы экотона горных ландшафтов оказались слабо разработанными. Хотя именно в горах экотоны хорошо прослеживаются при рассмотрении такого явления как высотная поясность.
Опыт полевых исследований в пределах Бокового хребта Северо-Западного Кавказа подтверждает это и позволяет проследить механизмы его проявления (Шальнев, 1971; Шальнев, Джанибекова, 1996; Шальнев, Юрин, 1997). Как правило, в горах выделяют два типа экотонов - ленточный и контурный (Шальнев, Нефедова, 2006). Ленточный проявляется чаще всего в виде переходных поясов на контрастных рубежах переходных зон от низкогорий к среднегорьям и от среднегорий к высокогорьям. В первом случае появление экотона связано с эффектом барьерного подножья и изменением качества увлажнения. На границе среднегорий и высокогорий проявляется эффект хионосферы связанный со снижением температуры июля до + 10°C и ниже. В связи с этим происходит увеличение криосферных дней со снежным покровом в течение года. Контурный тип экотона относится к внутриландшафтной дифференциации и представлен сочетанием морфологических единиц (фаций, урочищ) пограничных ландшафтов. Была разработана модель закономерностей формирования ленточных границ экотона, где важную роль играли климатогенные поля однородности (выделялись по суммарной радиации, радиационному балансу, средним годовым температурам воздуха и осадкам, температуре июля) и их смена по профилю рельефа гор. Именно границы этих полей служат условием формирования высотных геоботанических поясов экотона. Литогенные поля разнородности (морфоскульптуры и горные породы) определяют возникновение разнообразия поясов контурных экотонов. Контурные экотоны формируются в пределах ленточных экотонов. Их существование во многом определяется мезоформами рельефа. Например, древними и молодыми цирками, карами, висячими долинами, слаборасчлененными склонами. Кроме того, антропогенными факторами. Именно эти местности лучше всего осваивались человеком (вырубка леса, коши и т.д.) Так, в пределах Бокового хребта Западного Кавказа были выделены следующие основные виды контурных экотонов: цирковый, долинный, склоновой, хребтовый, ледниковый (Шальнев, Нефедова, 2006)К сожалению, стационарных исследований закономерностей вертикальной дифференциации компонентов ландшафтной среды на территории КЧР не проводилось. Исключением являются работы Тебердинского заповедника, проводимые на склоне хребта М. Хатипара в диапазоне высот 1400-2600м. над уровнем моря (Шальнев, Чикалин, 1968а, 1968б, Шальнев, 1971а, 1971б, Шальнев, Чикалин, 1971, Онищенко, 2002). В исследованиях принимала участие большая группа ученых-естественников под руководством А.А.Малышева. По всему профилю были установлены стационарные метеобудки и масляные осадкомеры на отметках 1400, 2050, 2500 и 2950 м над у.м. Наблюдения велись с 1966 по 1972 гг. Проводилась обработка метеоданных, что позволило рассчитать годовые показатели радиационного и теплового балансов, а также проследить особенности изменения градиентов метеоэлементов по высоте. В результате индивидуального подхода исследования проводились на трех ключевых участках, расположенных в пределах Бокового хребта Западного Кавказа, где были выделены следующие основные виды контурных экотонов: склоновый, хребтовый и ледниковый (Шальнев, Нефедова, 2006).Таким образом, понятие «географический экотон» отличается от биологического тем, что имеет более четкую пространственную обособленность и отражает «растянутость» границ перехода количественных и качественных изменений доминантных геоботанических поясов среднегорных и высокогорных ландшафтов Бокового хребта. Ленточный экотон формируется в общей структуре высотной поясности гор – ландшафтной и внутриландшафтной (геоботанических поясов). В зависимости от набора морфологических единиц «географического экотона» (местностей и урочищ) и соотношения по площади их лесных и луговых сообществ можно определять границу среднегорных лесных и высокогорных луговых ландшафтов, относя переходный пояс к тем или иным горным ландшафтам.
1.3. Ландшафтная экология
При рассмотрении этого вопроса, следует обратиться к истокам возникновения экологии и объединения её с географией. Зародившись в недрах биологии, биологическая экология (биоэкология) сформировалась как наука о закономерностях взаимодействия биологических систем с окружающей средой. Дальнейшее развитие экологии привело к образованию экологии человека и социальной экологии, то есть изучению взаимодействия со средой различных социальных систем (Кочуров, 1999).
У географии в связи с экологизацией науки появились благоприятные перспективы превратиться в науку синтетическую, опираясь прежде всего на необычайную ёмкость понятия “экология”. Впервые обратил внимание на роль географических исследований в решении экологических проблем академик В. Б. Сочава [1970]. Академик Н. П. Герасимов [1985] определил экологический подход в качестве общенаучного для всех конструктивных географических исследований. А. Г. Исаченко [1990] считает, что география больше других наук подготовлена к разработке экологической концепции.
С течением времени экологический принцип в географии стал завоевывать все более широкие позиции, и в первую очередь при исследовании биоты, биоэкосистем (биоцентрический подход) и ландшафтов (экология ландшафтов), также он более известен как геоцентрический подход. Другой важной разновидностью экологического принципа является антропоцентрический (антропоэкологический) подход, когда рассматривается взаимодействие человека с окружающей природной средой.
К. Тролль (Troll, 1939) обозначил науку, лежащую на рубеже географии (ландшафтоведения) и биологии (экологии) и назвал ее “ландшафтной экологией”. По определению А. Винка (Vink, 1983) главная задача ландшафтной экологии сводится к описанию и характеристике ландшафта в соответствии с его связями с биосферой и антропосферой.
В западно-европейской школе биогеографии ландшафтно-экологическое направление восходит к имени Г. Вальтера. Появляется и немецкая школа под руководством – Г. Хаазе и Э. Неефа (Нееф, 1974). Создателями современной концепции ландшафтной экологии считаются М. Годрон и Р. Форман (Forman, Godron, 1986). В нашей стране важную роль в развитии ландшафтной экологии сыграло развитие ландшафтной школы и ландшафтных методов при различных исследованиях биологических ресурсов. Таким образом, особенностью русской концепции ландшафтной экологии явилась ее заметная биоцентричность, с одной стороны, и использование достижений русской школы ландшафтоведения, с другой (Виноградов, 1994). Основоположник отечественной ландшафтной экологии В. Б. Сочава создал известный синтез экологии и географии сначала в Ботаническом институте (г. Ленинград) и затем, в особенности, в Институте географии (г. Иркутск) (Сочава, 1970, 1978). Полностью концепция ландшафтной экологии в России была сформулирована Б.В.Виноградовым (1994).
Биота непосредственно связана со средой, соответственно и группа хортобионтов привязана к среде обитания, растительность которой и определяет доминантные группы хортобионтов на определенном участке. Для формирования биоты определенного участка, в том числе и для хортобионтов, ведущими являются средоформирующие факторы. Основными такими факторами являются: местоположение (высотный пояс, экспозиция, крутизна склона), климатогенный (мезо- и микроклимат), трофический (почвы и растительность). Поэтому факторы среды не одинаково влияют на отдельные показатели структуры растительности, а соответственно, на численность, на видовой состав, на биомассу хортобионтов и их основные доминантные виды. Подобное соответствие рассматривалось на примере лесной (сосны и березы) и луговой растительности, потому что типы леса как природные системы в наибольшей мере обладают повышенной “чувствительностью” в отношении влияния факторов среды. Влияние же рельефа по всем характеристикам в сравнении с почвой существенно меньше. Это можно объяснить большей сопряженностью лесных и луговых ценозов с почвенными условиями, как прямодействующими.
1.4. Биотика ландшафта
В условиях все возрастающего антропогенного воздействия на биосферу происходят глобальные процессы потери ее генофонда, сокращения его разнообразия, площадей естественных ландшафтов и биоэкосистем с их «бесплатными» средоформирующими и ресурсовоспроизводящими функциями. Все это диктует необходимость поиска новых междисциплинарных подходов к познанию процессов формирования, развития и динамики природных и культурных ландшафтов с позиций оптимизации среды жизни человека, решения проблем сохранения ландшафтного и биологического разнообразия регионов.
К числу таких направлений можно отнести биотику ландшафта, под которой понимается научное направление, изучающее территориальное распределение биоты и биомов в рамках геосистем регионального и локального уровней и их роль в функционировании последних на основе ландшафтной парадигмы (Лиховид, Шальнев, Шкарлет и др., 2005). Это новое направление в ландшафтоведении, которое стало формироваться на кафедре физической географии Ставропольского госуниверситета с 2000 года в связи с тем, что исследование биотических компонентов ландшафта и в настоящее время остается слабо разработанным звеном теории ландшафтоведения. Более того, изучение этих вопросов происходит в рамках научного направления «Геофизика ландшафта», т.е. природе простейших форм движения материи. Перспективность этого направления определяется слабой изученностью биоты ландшафта – наиболее активного компонента, вовлекающего в круговорот неорганическое вещество и создающего биомассу (Исаченко, 1991).
Ведущей концепцией биотики ландшафта является биосферная концепция, которая сформировалась с учетом двух подходов:
- биосферного В.И. Вернадского, который рассматривал биосферу как глобальный биотоп Земли с большим разнообразием биострома;
- экобиосферного В.Н. Беклемишева, который изучал биосферу как экобиосистему планеты (биоценоз с биоценотическими и экологическими характеристиками), где разнообразие и совокупность биоэкосистем ландшафтов и их функциональные возможности обуславливают устойчивое состояние природной биосферной среды.
Методологической основой данного направления является ландшафтный подход, где природный ландшафт рассматривается как реальный объект исследования в качестве фокуса территориальной организации компонентов биосферы с позиций холизма (целостности взаимодействия абиотического, биотического и биокосного вещества в одном пространстве). Именно биота является тем «метахондрием» в генотипе ландшафтных геосистем, который определяет сущность взаимодействия абиотических, биотических и биокосных компонентов на определенном отрезке существования геосистемы.
В концептуальной основе биотики ландшафта лежит идея о полисистемной модели ландшафта (Шальнев, 1999, 2004). Она предусматривает выделение в рамках теоретического моделирования ландшафта трех подсистем: компонентной, морфологической и биоэкосистемной (биоценотической). В рамках этой модели биотика ландшафта находит возможности своей реализации на всех трех уровнях анализа.
В компонентной (топической) подсистеме изучаются вертикальные связи компонентов ландшафта, которые как в фокусе перекрещиваются в почвенном (биокосном) компоненте (рис. 1). Эти связи (взаимосвязи) обусловлены круговоротами субстрата, энергии и информации (СЭИ), в основе которых лежат физико-химические и биохимические процессы. Их особенности рождают имманентные свойства (состояние этих компонентов и режим их функционирования во времени в системе взаимосвязи), а также явление эмерджентности, т.е. природные условия, определяющие инвариантность (устойчивость) состояний компонентов и их связей. Биотический компонент в этой модели можно рассматривать с позиций количественных (видовое разнообразие) и качественно-количественных (продуктивность) характеристик. В топической или моносистемной модели разных видов геосистем изучаются вертикальные взаимосвязи всех компонентов, которые как в фокусе перекрещиваются в почвенном (биокосном) компоненте (рис. 1).Наиболее наглядно это можно проследить на примере самой малой геосистемы ландшафта – фации. Ведущей принцип анализа такой модели – равнозначность всех компонентов геосистемы.

Рис. 1. Топическая модель ранга фации (малой геосистемы) (Шальнев, 2004).
А – литогенное поле разнородности местности;
Б – климатогенное поле однородности геоботаниеского пояса.
Компоненты геосистемы:
1 – абиотические (горные породы, воздух, вода);
2 – биотические (растения, животное население, микроорганизмы);
3 - почвы (биокосные);
4 – вертикальные связи компонентов (на базе физико-химических и биохимических процессов);
5 – вход вещества, энергии и информации (ВЭИ);
6 – выход геосистем (качество природных условий, продуктивность).
Морфологическая (полисистемная) модель позволяет изучать закономерности внутриландшафтной дифференциации биоты и проводить территориальный анализ латеральных связей в системе катенных рядов. В их основе лежит понятие «литогенного поля разнородности», которое определяет пространственную структуру ландшафта и определяет при латеральных связях черты сходства и автономности фито- и зооценозов, формирует экотонную структуру границ.
В биоценотической подсистеме (подсистеме биоэкосистемного типа) возможно применение пространственно-временного анализа по принципу монодуализма с функциональной первичностью времени, что проявляется в сезонной динамике биоты, биохимических процессов и круговоротов СЭИ. Имманентная сущность связей в такой модели строится по принципу экоцентрических прямых и обратных связей, а эмерджентные свойства проявляются в поддержании инвариантных свойств качества природной среды жизни биоты. «Хозяин» модели (биота) испытывает не только воздействие компонентов окружающей среды, но и сам влияет на эти компоненты и качество среды. Причина – биохимические процессы и биотические источники энергии, определяющие особенности биогеохимических круговоротов вещества в пределах биоэкосистемы. В таком качестве набор биоэкосистем (в ранге фаций) представляет собой «анатомию» и «физиологию» ландшафтной структуры, которая формирует и регулирует природную среду не только ландшафта, но и биосферы в целом (рис. 2).
Биоэкосистемы ландшафта, их имманентные и эмерджентные свойства испытывают внешние воздействия более крупных морфологических единиц – местностей и высотных геоботанических поясов. Например, климатогенное поле однородности высотного геоботанического пояса определяет инвариантное проявление совокупности элементов биотического компонента (как фито, так и зоо). Его влияние проявляется через гидротермические условия, сезонная динамика которых определяет и сезонную динамику элементов биоэкосистемы. Литогенное поле разнородности ландшафта вносит свои поправки в гидротермические условия и определяет территориальную мозаику биотических сообществ. Например, в поясе хвойных лесов Северо-Западного Кавказа чаще всего на склонах северной экспозиции растут пихтовые леса, южной – сосновые.

Рис. 2. Биоценотическая модель фации (биоэкосистемы) (Шальнев, 2004).
1 – биоценотическая модель.
Ее компоненты:
2 – абиотические (горные породы, воздух, вода);
3 - растительность;
4 – животное население;
5 - микроорганизмы;
6 – почвы.
Подсистема:
7 - “среда”.
Связи геосистемы:
8 – влияние компонентов среды на “хозяина”
9 – обратная связь “хозяина”;
10 – биогеохимический круговорот.
Внешние геосистемы:
А – литогенное поле разнородности ”местности”;
Б – климатогенное поле однородности геоботанического пояса;
11, 12 – прямые и обратные связи с внешней средой.
13 – вход ВЭИ;
14 – выход геосистемы (качество окружающей среды как части экобиосферы Земли).
В традиционных ландшафтных исследованиях, и в настоящее время, основное внимание уделяется растительности и животному населению с позиций компонентного анализа с использованием описательных методов. Внутриландшафтная дифференциация биотического компонента обычно рассматривается на примере изучения биогеоценозов. Подобный же опыт изучения животного населения фактически отсутствует. С этой проблемой мы столкнулись при изучении ландшафтов Северного Кавказа.
В свою очередь результаты этих круговоротов (наращивание биомассы, ее отмирание и утилизация) оказывают большое влияние на компоненты окружающей среды биоценотической модели (особенно почвенные), а также, как следствие, на само качество окружающей среды не только биоэкосистемного уровня, но и ландшафтного и в целом экосферного биосферы Земли. Таким образом, биоэкосистемы – это набор “атомарного уровня” ландшафтных структур, формирующих и регулирующих качество природной среды ландшафта и биосферы.
Биоэкосистемы ландшафта, их имманентные и эмерджентные свойства испытывают значительные внешние воздействия более крупных геосистем (ранга местности и высотного геоботанического пояса). Например, “климатогенного поля однородности” высотного геоботанического пояса среднегорий, которое определяет инвариантное проявление совокупности элементов биотического компонента (как фито-, так и зоо-). Его влияние проявляется через гидротермические условия, сезонная динамика которых определяет и сезонную динамику элементов биокомпонента. “Литогенное поле разнородности” ландшафта, геоботанического пояса, местности (абсолютная и относительная высота, горные породы, мезо- и микроформы рельефа, экспозиция и др.) вносит свои поправки в гидротермические условия и, следовательно, в суженный инвариант биотического компонента (например, в хвойном поясе → сосновые леса занимают склоны южной экспозиции а пихтовые леса - склоны северной экспозиции. Также у них верхняя граница произрастания разная – у сосновых лесов – 2300-2355, а у пихтовых – 1950-2000 м. над уровнем моря).
Глава II. Ведущие природные факторы внутриландшафтной дифференциации беспозвоночных высокогорий
2.1. Геоморфологический фактор
Большой Кавказ в тектоническом отношении является эпигеосинклинальной геотектурой. Он имеет резко выраженное асиммитричное строение, обусловленное системой крутых сбросов и надвигов и подразделяется на два типа тектонических структур – блоковые или глыбовые хребты на древнепалеозойском фундаменте и депрессии. К блоковым хребтам относятся Главный, Боковой и Передовой, протянувшиеся в субширотном направлении. К депрессиям относятся Южно-Юрская, сформированная осадочными отложениями нижнеюрского возраста, и Архызско-Загеданская средне- и верхнеюрского возраста. По тектоническим разломам формировались эрозионно-тектонические долины, такие как Теберда, София, Кизгич и другие (Герасимов, Лилиенберг, 1984).
Исследования проходили на территории Западного Кавказа по трем высокогорным ландшафтам, в которых наиболее четко прослеживается смена гидротермических условий в долготном направлении. Данный район выделяется как по амплитуде неотектонических поднятий и характеру структур, так и по характеру растительности и климатическим характеристикам (Милановский, Хаин, 1963; Белановская, 1990; Ефремов и др., 2001). Горный рельеф изучаемой территории формируется под воздействием морозного выветривания, речной эрозии, склоновой денудации и ледниковой деятельности, ограниченной в пространстве ареалом распространения ледников. На характер проявления этих процессов в значительной степени влияют реликты древнеоползневых и сейсмогенных форм. В масштабе региона они хорошо развиты, чему способствует густая сеть разрывов и тектонических трещин, разбивающих массивы на блоки разных порядков (Черных, 1991).
Морфометрическая и морфологическая характеристика исследуемых морфоструктур Западного Кавказа отличается друг от друга.
В основе геоморфологического фактора лежит геологическое строение. Горст-антиклинорий Главного хребта, развившийся в пределах северной краевой зоны геосинклинали Большого Кавказа, играет в современной структуре Западного Кавказа роль осевого поднятия. Он сложен кристаллическими сланцами и гнейсами нижнего и среднего палеозоя, на значительных пространствах замещенными гранитоидами средне- и верхнепалеозойского возраста (Герасимов, Лилиенберг, 1984).
Главный Кавказский хребет представляет собой эрозионно-тектонические горы и является водораздельным хребтом, в рельефе которого преобладают карлинги с крутыми вершинами. Общая длина Главного хребта 1385км, при средней высоте 2900м. Максимальная высота хребта 5182м (г. Шхара). Относительные высоты Главного хребта в пределах Западного Кавказа повышаются с запада на восток. Основными вершинами являются: Фишт (2868м), Чугуш (3238м), Цахвоа (3345м), Пшиш (3790м), Эрцахо (3910м), Домбай-Ёльген (4046м), Далар (3988м) (Ефремов и др., 2001, с. 56-57). Сложен он толщами протерозойских и палеозойских кристаллических сланцев и крупными интрузиями гранитоидов. Горст-антиклинорий хребта осложнен системой глубоких разломов, разделяющих его на ряд кулисообразных блоков (Софийский, Тебердинский и другие) (Шальнев, Джанибекова, 1996).
Граница между Главным и Боковым хребтами не везде четко выражена, что обусловлено сложным взаимодействием структурных блоков (покровных пластин). Наиболее определенно раздел между ними наблюдается на северной стороне Главного хребта. Он проходит по долинам рек, совпадающим со структурной зоной, называемой Южно-Юрской депрессией. Ее Домбайский участок представлен абсолютными отметками 1800-1900м над уровнем моря. Это троговая долина, у которой склоны сложены метаморфическими породами палеозоя, а днище заполнено четвертичными моренными, флювиогляциальными и аллювиальными отложениями.
Боковой хребет расположен к северу от Главного на расстоянии от 2км. до 25км. и южнее Передового на расстоянии 8-22км. В среднем, общая длина Бокового хребта 662км, при средней высоте 2310м. Основные вершины хребта: София (3687м), Большая марка (3758м), Даут (3748м), Курша (3870м), Куршоу (3690м) и т.д. Он образован выступом герцинского фундамента, сложенным кристаллическими сланцами и гнейсами нижнего и среднего палеозоя и гранитоидами позднего палеозоя. Особенностью этой зоны является наличие в ней крупных интрузий габбро и ультраосновных пород (серпентиниты), кварцевых диоритов, гранитов, а также эффузивных образований. Однако участие в тех или иных геологических формаций в строении центральной зоны неодинаково (Ефремов и др., 2000).
От Бокового и Главного хребтов Передовой хребет отделяется на западе Архызско-Загеданской внутригорной эрозионно-тектонической продольной депрессией, сложенной нижнеюрскими отложениями. Архызский новейший грабен представляет собой северное ответвление от основного желоба шовно-депрессионной зоны. Он разделен на блоки меридиональными разломами и речными долинами.
К северу от Бокового хребта, на участке от р. Белой на западе до р. Урух на востоке, протянулся Передовой хребет, являющийся самостоятельной орографической единицей. Общая длина хребта 314км, средняя высота 2970м. Основные его вершины Кынгыр-Чад (3543м), Кенделляр-Ляр (3416м), Кызыл-Кол (3638м). Он сложен метаморфическими и осадочными породами палеозоя. Особенно сложен состав горных пород в районе хребта Абишира-Ахуба, где в большей мере распространены среднепалеозойские сильно метаморфизованные лавы, а также пестроокрашенные песчаники, алевролиты, аргиллиты и другие породы (Михеев, Снежко, Сафронов, 1962).
Основными видами морфоструктур, формирующихся в пределах геотектуры Большого Кавказа могут называться:
1) водораздельные хребты субширотных склонов северной экспозиции высокогорий Главного Кавказского хребта с карлингами и перевалами.
2) троговые долины Южно-Юрской субширотной депрессии (Домбайская, Софийская), которая отделяет Главный хребет от Бокового хребта.
3) троговые меридиональные эрозионно-тектонические долины рек Теберды, Кизгича, Софии и другие в пределах Бокового хребта.
4) южные крутые и обрывистые склоны Передового хребта и Архызско-Загеданская депрессия с субширотными долинами рек, отделяющая Боковой хребет от Передового хребта.
5) эрозионно-тектонические долины рек меридионального простирания, разделяющие блоки Передового хребта.
6) склоны осевых хребтов разных экспозиций высокогорий Бокового хребта и Передового хребтов.
В пределах этих типов морфоструктур формируются морфоскульптуры, среди которых широко распространены формы ледникового происхождения: цирки, кары, карлинги, морены – боковые и конечные, озерные четки и др. Достаточно широко распространены эрозионно-аккумулятивные формы рельефа – коллювиальные склоны притоков вторичных и третичных долин, речные террасы, эрозионные борозды, конуса выносов. Встречаются коллювиально-пролювиальные формы рельефа – конуса выноса селевых потоков и карстовые формы рельефа типичные для Передового хребта, например, карстовые озера.
2.2. Климатогенный фактор
На климатические условия Западного Кавказа влияют географическое положение, особенности рельефа и циркуляция атмосферы. Участок расположен на юге умеренных широт на границе с субтропическими широтами (Асланикашвили, Мамулян, 1990). Климат влажный, относительно солнечный и теплый, так как осуществляется средиземноморский циклогенез. В холодное полугодие количество осадков велико. На высоте около 2000м за год выпадает 2600 мм. (Темникова, 1959). На территории Западного Кавказа преобладает юго-западный перенос общей циркуляции. Он для северных склонов Передового хребта и Южно-Юрской депрессии приносит основную массу осадков. В годовом и суточном ходе различных метеорологических элементов происходят резкие изменения, определяемые взаимодействием свободной атмосферы с увеличением высоты. Характер изменения температуры с высотой не зависит от сезона: количество осадков растет с высотой (Кавказ, 1966). В общем, для высокогорных районов характерен высокий приток суммарной солнечной радиации. Но от среднегорий к высокогорьям наблюдается уменьшение радиационного баланса, в связи с повышением эффективного излучения с высотой. Наблюдается общее возрастание засушливости с запада на восток.
Имеющаяся на территории исследования метеостанция «Клухорский перевал» на высоте 2037м над у.м., располагает данными, которые помогут оценить изменения климатических параметров за с 1960 по 2004 гг. (таблица 1). Для анализа была выбрана эта метеостанция потому, что она расположена в типичной зоне экотона субальпики. Нами при изучении участков исследования было проанализировано изменение годовых температур в период по данным метеостанции.
Таблица 1.
Температура воздуха за период 1960-2004 гг. по метеостанции «Клухорский перевал», ºС (Братков, 2005)
| 196 1960-2004гг. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | Год |
|
Tmin. |
-10,1 | -9,6 | -6,2 | 0,0 | 4,5 | 8,4 | 10,6 | 9,5 | 6,3 | 0,5 | -4,3 | -7,4 | 2,1 |
|
Tmax. |
-0,2 | 4,2 | 2,8 | 8,5 | 9,5 | 14,4 | 17,3 | 16,3 | 12,5 | 10,1 | 5,7 | 3,9 | 5,9 |
|
Tср. |
-5,0 | -4,6 | -1,8 | 3,0 | 7,0 | 10,3 | 13,2 | 12,7 | 6,3 | 5,3 | 0,6 | -2,5 | 4,0 |
Минимальная температура воздуха составила 2,1ºС в 1992 г., а максимальная – 5,9ºС в 1971 г. Если сравнивать тенденции изменения температуры по периодам, то c 1960-х гг. до 1990-х гг. колебания осуществлялись в диапазоне 4ºС. В начале 90-х гг. диапазон колебаний средних температур составил 3ºС, а с конца 90-х гг. снова наблюдается повышение показателей до 4,2ºС-4,3ºС. Отмечается несовпадение данных по многолетним наблюдениям: при увеличении температуры за указанный период наблюдений выявляется, наоборот, падение температуры. Данный факт можно объяснить тем, что в начале 90-х годов отмечались минимальные температуры за весь анализируемый срок наблюдений. Кроме того, примерно до середины 70-х годов амплитуда колебания температуры воздуха была гораздо более существенна, по сравнению со следующим периодом (Братков и др., 2005).
Отрицательные минимальные температуры характерны для пяти месяцев в году, начиная уже с ноября, и, соответственно, самая низкая температура бывает в январе -10,1ºС. Отрицательные максимальные температуры характерны только для января. В последующие месяцы рост показателей осуществляется примерно с разницей в два раза. Положительные максимальные температуры актуальны для трех летних месяцев и колеблются от 14,4ºС до 17,3ºС.
Для средних температур динамика показателей изменяется примерно в два раза. Отрицательные средние показатели характерны для четырех месяцев - декабрь, январь, февраль и даже март, причем низкий показатель встречается в январе, а немного «повыше», соответственно, в марте. Такие температурные вариации сокращают вегетационный период развития растительности и активный образ жизни беспозвоночных. Максимальные показатели средних температур выделяются в летние месяцы и диапазон колебаний составляет от 10,3ºС до 13,2ºС. Именно этот промежуток времени и характеризуется активизацией процессов жизнедеятельности беспозвоночных.
Наглядно динамику общих макроклиматических показателей можно проанализировать на примере метеоэлементов хребта Малая Хатипара.
Значительные колебания относительных высот в пределах хребта обусловили формирование вертикальных различий климата, растительности, почв и животного мира. При рассмотрении основных показателей метеоэлементов климата (таблица 2) —радиационного баланса, затрат тепла на испарение, индекса сухости, испаряемости — отмечается общая тенденция снижения показателей с высотой. Средние июльские температуры снижаются на 0,5° на каждые 100м, средние годовые - на 0,4°, радиационный баланс — на 0,7 ккал/см2, затраты тепла на испарение — на 0,19 ккал/см2, затраты тепла на нагревание - на 0,5 ккал/см2 на 100м. В то же время количество осадков и величина коэффициента увлажнения растут с высотой. Количество осадков возрастает на 64 мм на каждые 100 м, величина коэффициента увлажнения — на 0,4.
Составляющие теплового баланса с высотой меняются одинаково. Затраты тепла на испарение по всему профилю хребта изменяются мало, чего нельзя сказать о затратах тепла на нагревание. Годовые величины последних с высотой быстро уменьшаются. В данном случае показателен коэффициент отношения затрат тепла на испарение к затратам тепла на нагревание (LE/P). В долине Теберды величины LE и Р почти одинаковы, и коэффициент равняется 1,27. В пределах лугового пояса его величины возрастают до 2,0—2,13.
Уменьшение затрат тепла на нагревание с высотой сказывается на температурном режиме воздуха и характере испаряемости. В поясе луговых ассоциаций средние годовые температуры уже ниже нуля. Величины испаряемости не превышают 300мм, поэтому коэффициент увлажнения растет с 1,3 в долине Теберды до 4,8—5,8 в поясе луговых ассоциаций. Подобных значений коэффициента увлажнения у природных зон равнин умеренных широт не наблюдается (Шальнев, 1973).
При сравнении показателей таблицы 2 от подножия (1340м) к субальпийским лугам на высоте 2500м. над у.м., выявлено, что основные показатели метеоэлементов весьма высоки для станции 1, а на второй станции наблюдается спад показателей. От этой станции вполне упорядоченно возрастают метеоэлементы к типичной субальпике. Максимальные показатели температур характерны для высоты в 1340м, потом резкое падение для июльских температур на 4,3ºС, а для годовой – 1,9ºС и более плавное понижение, в результате которого годовая температура и температура воздуха в июле понижается на 0,6-1,0ºС, а годовая температура в диапазоне высот 2350-2500м даже понижается на 1,9ºС. При такой динамике температур минимальное количество осадков выпадает в хвойно-широколиственных лесах – 763 мм, затем повышается количество осадков, причем на 479 мм и на верхней границе пихтово-сосновых лесов составляет 1410 мм. А к субальпийским лугам количество осадков увеличивается плавно – на 84-168 мм. Но при таком росте количества осадков, влажность воздуха с высотой уменьшается: минимальная на станции 4 (68 мм), а максимальная – на станции 1 (76 мм). Радиационный баланс с высотой уменьшается от 38,0 ккал/см2 до 30,1 ккал/см2, причем разница между первыми двумя станциями составляет 6,9 ккал/см2. Расходная часть радиационного баланса, которая тратится на затраты тепла на испарение (LE) и турбулентный поток тепла в воздух (P). Вполне последовательно понижается показатели на испарение от 1340 м до 2500 м над у.м., с разницей 0,1 ккал/см2. Показатели P сначала понижаются на 6,3 ккал/см2, затем не изменяются на уровне высот 2050 м. и 2350 м и составляют 10,2 ккал/см2. На станции 4 показатель турбулентного потока тепла в воздух составляет всего лишь 10,2 ккал/см2. Затраты на испаряемость закономерно снижаются с 430 мм до 312 мм., это объясняется тем, что луговые ассоциации являются «открытым» участком, лишенным древесной и кустарниковой растительности. Соответственно, расходная часть радиационного баланса от леса к лугу возрастает.
При сравнении березовых криволесий и сосновых редколесий с типично субальпийскими лугами при разнице высот в 150м наблюдается снижение средней июльской и годовой температур на 1º и 1,5°, соответственно, и уменьшается влажность воздуха на 6%. При этом, от станции 3 к станции 4 возрастает количество осадков на 84 мм., то и радиационный баланс, затраты тепла на испарение и турбулентный поток тепла в воздух также возрастает, но незначительно на 0,1 ккал/см2 , 0,7 ккал/см2 и 0,2 ккал/см2 , соответственно. Испаряемость уменьшается на 10 мм, а значит и коэффициент сухости – на 0,3. Коэффициент увлажнения, наоборот, возрастает на 0,4. Так как, возрастание радиационного баланса параллельно ведет к нарастанию и эффективного излучения.
Таблица 2.
Изменение основных показателей метеоэлементов климата
по восточному профилю хребта Малая Хатипара (Шальнев, 1968-1975).
| Станции |
Абс. высоты в м. |
Температура воздуха |
Осадки за год в мм. |
Влажность воздуха |
Ккал./см² | Кс | Кв | LE/P | Испаряе-мость в мм. | ||||
| июль |
годо- вая |
% | мб. | R | LE | P | |||||||
| Станция 1 (пояс хвойно-широколиственных лесов) | 1340 | 15,6 | 6,3 | 763 | 70 | 6,7 | 38,0 | 21,3 | 16,7 | 0,83 | 1,3 | 1,27 | 430 |
| Станция 2 (верхняя граница пихтово-сосновых лесов) | 2050 | 11,2 | 3,4 | 1242 | 76 | 5,9 | 31,1 | 20,7 | 10,4 | 0,42 | 3 3,55 | 1,98 | 350 |
| Станция 3 (березовое криволесье, сосновые редколесья и субальпийские луга) | 2350 | 10,6 | 2,7 | 1410 | 74 | 5,5 | 31,0 | 20,6 | 10,4 | 0,37 | 4,4 | 1,96 | 322 |
| Станция 4 (субальпийские луга) | 2500 | 9,6 | 0,8 | 1494 | 68 | 4,4 | 30,1 | 19,9 | 10,2 | 0,34 | 4,8 | 1,95 | 312 |
Примечание: R – радиационный баланс, LE – затраты тепла на испарение, P – турбулентный поток тепла в воздух, Кс – коэффициент сухости, Кв – коэффициент увлажнения.
При сравнении средних температурных показателей за 44 года (Братков, 2005) и за 7 лет (Шальнев, 1973) выявлено, что за многолетний период изменения составляли около 4ºС на уровне 2037 м, а за семилетний промежуток на высотах от 1340м до 2350м над у.м. диапазон колебаний составлял 0,3-0,4ºС. Среднегодовая температура на лугу в типичной субальпике (2500м) достигла 2ºС. За более короткий временной промежуток времени градиент колебаний будет более сглаженный, чем за почти полувековой период, причем показатели, безусловно, усреднены, а общий анализ дан выше.
Еще одним показателем, характеризующим климатическую обстановку Западного Кавказа является количество осадков. Также можно проанализировать изменение величины годовых осадков на метеостанции «Клухорскиий перевал» за период 1960-2004 гг. (таблица 3), для получения многолетних показателей. По сравнению с предшествующим периодом годовое количество осадков увеличилось на 23 мм, что при средней величине около 1800 мм не существенно, однако сезонные изменения довольно весомы. Заметна хорошо выраженная тенденция увеличения осадков в холодный период при их сокращении в теплый период года. При этом в процентном исчислении, например, в январе количество осадков увеличилось на 42%, тогда как в мае они уменьшились на 35%.
Таблица 3.
Осадки за период 1960-2004 гг. по метеостанции «Клухорский перевал»,
в мм (Братков, 2005).
| 1960-2004гг. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | Год |
|
Осадкиmin. |
10 | 3 | 4 | 81 | 22 | 51 | 32 | 28 | 39 | 24 | 21 | 10 | 1301 |
|
Осадкиmax. |
498 | 305 | 378 | 342 | 290 | 346 | 277 | 259 | 425 | 563 | 439 | 440 | 2377 |
|
Осадкиср. |
151 | 104 | 119 | 164 | 144 | 149 | 137 | 142 | 1581 | 196 | 177 | 163 | 1798 |
Минимальное количество осадков составляет 1301 мм в 1984 г., а максимальное – 2377 мм в 2001 г. Если сравнивать тенденции изменения осадков последовательно, то с 1960-х гг. их величина составляла 1718 мм, и до 1980-х гг. возрастает – 1763 мм, а в 80-ые гг. резко уменьшается – до 1575 мм. Но от 90 гг. к 2004 г. опять увеличивается среднегодовое количество осадков и составляет уже 2102 мм. Величина гидротермического коэффициента за период 1960-2004 гг. составила 3,82, т.е. уменьшилась с 4,6. Причем, гидротермический коэффициент представляет собой отношение суммы осадков за период с температурой выше +10º к сумме температур за тот же период. Величина коэффициента увлажнения, наоборот, несколько увеличилась: 3,52 вместо 3,40. А коэффициент увлажнения представляет собой отношение количества осадков к испаряемости за тот же период (Братков и др., 2005).
Самое минимальное количество осадков, исходя из таблицы 3, характерно для февраля и марта, затем резко возрастает в апреле и в летние месяцы колеблется от 51 до 28. Следующий пик достаточного увлажнения приходится на сентябрь. К январю наблюдается уменьшение количества осадков. При этом наибольший показатель осадков из критерия «Осадкиmin» отмечен в апреле – 81 мм, а минимальный – в феврале – 3 мм. Максимальное количество осадков изменяется вполне закономерно: значительно для осеннее-зимнего периода от 425 мм в сентябре до 498 мм в январе и максимум приходится на октябрь. В весеннее-летний период осадки уменьшаются: от 378 мм в марте (максимум) до 259 мм в августе (минимум).
Среднее количество осадков составляет для полувекового промежутка времени 1775 мм., где наименьшее количество 104 мм выделено в феврале, а наибольшее – 1581 мм в сентябре. В целом, в осеннее-зимние месяцы, с сентября, количество осадков уменьшается и в феврале достигает 104 мм. Весной их количество увеличивается и в летний сезон несколько уменьшается.
Основными особенностями ветрового режима Западного Кавказа является наличие горно-долинной циркуляции с направлением ветра вдоль долин и преобладанием ветров западных направлений. Как правило, наибольшие скорости ветра здесь отмечаются в холодной период, а наименьшие – в теплый. Вероятность ветров скоростью более 10 м/с повышается зимой - 40 % (Гельмгольц, 1963).
Горно-долинные ветры охватывают долины от их верховьев до предгорий. Развиты они в течение всего года, но наиболее интенсивно в теплый период при антициклональных типах погоды, когда барические градиенты ослаблены. Днем ветер имеет направление вверх по долинам и склонам – долинный ветер, а ночью вниз по долинам – горный ветер. На северном склоне дневной долинный ветер имеет северное, северо-восточное направление, а ночной, горный ветер – южное и юго-западное направление. Также повсеместно наблюдаются фены, которые обусловлены, в основном, циклонической деятельностью. Их возникновение связано с происхождением циклонов средиземноморского происхождения (Тайлаков, 1967).
Помимо макроклимата в горах, приводящего к дифференциации природных условий и формированию биоклиматических комплексов — геоботанических высотных поясов, имеются и микроклиматические особенности, воздействующие на горизонтальную дифференциацию. Это экспозиция и крутизна склонов. Экспозиция склона в первую очередь влияет на обеспеченность теплом природных комплексов. При одинаковом количестве осадков неравномерное распределение тепла определяет различия в водном режиме склонов, что сказывается на особенностях биоценозов (Шальнев, 1971). Поэтому в пределах одного и того же высотного пояса «в одинаковых" условиях общего климата на близких друг к другу участках могут создаваться местные вариации климата, что влияет на детали структуры ландшафтной поясности» (Щукина, I960, стр. 17).
Таблица 4.
Распределение годовых величин теплового и радиационного балансов в геоботаническом высотном поясе березовых криволесий, сосновых редколесий и субальпийских лугов, в зависимости от экспозиции склона, ккал/см² (Шальнев, 1973)
| Станции | R | LE | P | Кэ | |
| Северная | 29.6 | 18,9 | 10.7 | 0,4 | |
| Южная | 38,5 | 23.6 | 14,9 | 3,3 | |
| Восточная | 31,3 | 20.1 | 11.2 | 1.8 | |
Примечание: R – радиационный баланс, LE – затраты тепла на испарение, P – турбулентный поток тепла в воздух, Кэ – коэффициент экспозиции.
Примером формирования местных вариаций климата на склонах разных экспозиций, на близких друг к другу участках, могут служить данные, приведенные в таблице 4. Станции располагались в пределах луговых ассоциаций переходного геоботанического пояса березовых криволесий и сосновых редколесий с фрагментами субальпийских лугов. Первая станция (северная) занимала пригребневые участки склона северо-северо-восточной экспозиции и крутизной 16—18° с разнотравно-злаковыми лугами. Вторая станция (южная) располагалась на склонах юго-юго-восточной экспозиции с крутизной 30°, с разнотравно-пестрокостровыми фитоценозами. Восточная станция занимала крутые (20°) склоны верхней части древнего цирка восточной экспозиции с вейниково-злаково-разнотравной pacтительностью.
Для иллюстрации роли экспозиции в пределах одного высотного геоботанического пояса очень показателен коэффициент экспозиции (Кэ), который рассчитывался В.А. Шальневым пo формуле Кэ = P×A / < K×R, где А—азимут склона, < K—крутизна склона. На склоне северной экспозиции при А = 15° коэффициент равняется 0,4. На склоне южной экспозиции при А=165˚ достигает 2,3. Для склонов восточной экспозиции при А =105˚ коэффициент составляет величину 1,8 (Шальнев, 1973).
2.3. Ландшафтный фактор
Высотный фактор определяет вертикальную дифференциацию климатических поясов, а геоморфологический фактор формирует внутрипоясные различия, что приводит к выделению остальных морфологических единиц, например, местностей – это цирки, склоны долин второго и третьего порядков, висячие долины и т.д. Важность геоморфологического фактора заключается в обособлении и смене растительности в высокогорьях. Также изменение с высотой климатических условий тоже определяет дифференциацию растительного и почвенного покрова и, как следствие, формирование высотных геоботанических поясов. Это влияние, а также соотношение ландшафтных и геоботанических поясов, в том числе и поясов экотона, можно рассмотреть также на примере Тебердинского заповедника (левого борта долины реки Теберды и хребта Малая Хатипара) (рис.3).

Рис. 3. Ландшафтные и геоботанические пояса Тебердинского заповедника
(Шальнев, Нефедова, 2006).
Условные знаки:
Границы: 1 – ландшафтов, 2 – геоботанических поясов.
R – радиационный баланс, ккал/см2 ; t1 – средняя годовая температура; t2 – температура июля; r– годовая сумма осадков.
Геоботанические пояса ландшафта хвойных лесов троговых долин среднегорий:
I – доминантный хвойных лесов склонов трогов;
II – экотона хвойно-широколиственных лесов днища трога;
III – экотона сосновых редколесий, березовых криволесий и высокотравных субальпийских лугов склонов трогов.
Геоботанические пояса ландшафтов высокогорных лугов:
IV – доминантный субальпийский;
V – доминантный альпийский;
VI – экотон субнивальный.
В пределах среднегорий формируется ландшафт хвойных лесов меридиональных троговых долин, для которых типичны свои геоботанические пояса. На днище троговой долины реки Теберды формируется переходный пояс смешанных лесов. Основные компоненты колеблются в пределах: R = 37-38 ккал/см2, t1 = 6,3ºС, t2 = 15,1-15,6ºС, r = 760-850 мм, что позволяет произрастать не только хвойным лесам, но и буковым лесам. На склонах трогов формируется доминантный пояс хвойных лесов. Показатели метеоэлементов изменяются: R = 37-31,5 ккал/см2, t1 = 6,0-3,0ºС, t2 = 15,0-11,2ºС, r = 850-1250 мм, благодаря чему в зависимости от фактора экспозиции произрастают сосново-пихтовые леса. При дальнейшем увеличении высоты формируется переходный пояс сосновых редколесий и березовых криволесий к субальпийским лугам. Здесь основные компоненты метеоэлементов составляют: R = 31 ккал/см2, t1 = 2,7ºС, t2 = 9,6ºС на южных склонах и 9,9ºС – на северных, r = 1400 мм. Данные климатические условия показывают постепенную зону перехода от лесных ассоциаций к высокогорным луговым. Причем температура достигает здесь критической отметки в 10ºС, которая не является благоприятной для произрастания древесного и кустарникового ярусов. Если обратиться к разнице высот границ, то на южных склонах граница экотона занимает более высокое положение, а на северных как бы приостанавливается.
В пределах высокогорного лугового ландшафта хребта Малая Хатипара также выделяются геоботанические пояса. Доминантный субальпийских лугов характеризуется следующими показателями метеоэлементов: R = 30,5 ккал/см2, t1 = 0,8ºС, t2 = 9,0-9,6ºС, r = 1500 мм. Средняя годовая температура здесь близка к 0ºС, температура июля ниже 10ºС. Данный пояс расположен до отметок 2500-2600 м над у.м. Выше, где климатические условия становятся более суровыми, размещается доминантный геоботанический пояс альпийских лугов. Показатели его основных метеоэлементов: R = 28 ккал/см2, t1 = -0,7ºС, t2 = 8,5ºС, r = 1600 мм.
В растительном покрове преобладают низкорослые виды, а также мхи. При дальнейшем увеличении высоты формируется разорванный маломощный растительный покров представляет собой переходный геоботанический пояс – субнивальный. Здесь альпийские луга встречаются пятнами, много осыпей и снежников. Последние сохраняются и летом (Шальнев, Нефедова, 2006).
Объектом изучения явилась зона экотона горных ландшафтов среднегорий и высокогорий. Высокогорные пояса экотона формируются посредством фактора высотной поясности и наложением на него геоморфологического фактора (наличие морфоскульптур). А в силу того, что группа хортобионтов является индикационным фактором растительности высокогорных поясов и лугов, то следует рассмотреть ландшафты исследуемого участка Западного Кавказа. Ландшафтный подход использовался для изучения ландшафтов среднегорий и высокогорий – это Главный Кавказский, Передовой и Боковой с долинами. Они входят в Лабино-Тебердинский округ, к которому относится верхний участок ландшафтного высотного пояса среднегорий темно-хвойных лесов и нижняя часть ландшафтного высотного пояса высокогорий субальпийских и альпийских лугов (Шальнев, Джанибекова, 1996). Составлены ландшафтные карты, из которых выбраны те территории, где нами проводился эксперимент – это ландшафты Домбайской депрессии (доминантный пояс хвойных лесов и переходный пояс хвойно-широколиственных лесов), среднегорный и высокогорный ландшафты хребта Малая Хатипара и южный склон Передового хребта в районе среднегорий и депрессии.
Ландшафты Лабино-Тебердинского округа.
I. Ландшафтный высотный пояс среднегорий смешанных и хвойных лесов. Выделяются виды ландшафтов:
1. Внутригорных эрозионно-тектонических депрессий Главного хребта (Домбайская), сложенных нижнеюрскими осадочными толщами и коренными породами палеозоя, со смешанными и хвойными лесами на бурых остаточно-оподзоленных горно-лесных почвах (участок долины реки Птыш). Климат умеренно теплый с годовыми величинами суммарной радиации (Q) 112-127 ккал/см2, радиационного баланса (R) 35-40 ккал/см2, средними годовыми температурами воздуха 4,0-6,5ºС и суммами осадков 900-1200 мм. Прослеживаются геоботанические пояса:
а) переходный горных лугов с куртинами лесной растительности (ольшаники, березовые, кленовые, пихтовые) на горно-луговых и горно-лесных почвах, занимающий днища Домбайской поляны и речных долин Алибека и Домбай-Ульгена, сложенные аллювиальными и моренными отложениями. Вероятность участия антропогенного фактора в формировании;
б) переходный смешанных (буково-пихтовых) лесов на бурых горно-лесных почвах верхней трети коллювиальных склонов троговых долин;
в) хвойных (пихтовых) лесов на грубоскелетных бурых горно-лесных почвах верхней трети коллювиальных склонов троговых долин;
г) переходнвй пояс березовых криволесий, пихтовых редколесий с фрагментами высокотравных субальпийских лугов на горно-кустарниковых и горно-луговых почвах (Шальнев, 2005).
2. Меридиональных троговых долин Главного хребта, сложенных серыми гранитами палеозоя (хребет Малая Хатипара). Четвертичные отложения представлены коллювием и ледниковыми отложениями. Климат умеренно теплый с годовыми величинами Q = 125-130 ккал/см2, R = 30-35 ккал/см2, средними годовыми температурами воздуха 4,5-5,5ºС и суммами осадков 800-2000 мм. Выделяются геоботанические пояса:
а) переходный пояс смешанных (пихтово-буковых) лесов на бурых горно-лесных и бурых оподзоленных почвах днищ троговых долин;
б) доминантный пояс пихтовых и сосново-пихтовых лесов на бурых горно-лесных почвах склонов троговых долин;
в) переходный пояс березовых криволесий, сосновых (пихтовых) редколесий с фрагментами субальпийских лугов плечей трогов.
II. Ландшафтный высотный пояс высокогорий субальпийских и альпийских лугов.
Выделяются виды ландшафтов:
6. Ландшафты Передового хребта, сложенные метаморфическимим и осадочными породами палеозоя с реликтовыми ледниковыми формами рельефа, каровыми и карстовыми озерами (южный склон хребта Абишира-Ахуба). Климат умеренно холодный и холодный. Выделяются геоботанические пояса:
а) доминантный субальпийских лугов с рододендроном кавказским на бурых дерновых горно-луговых и бурых торфянистных горно-кустарниковых почвах;
б) доминантный альпийских лугов на горно-луговых почвах;
в) переходный субнивальный со скальными выходами пород, пятнами альпийских лугов и мохово-лишайниковыми пустошами на фрагментарных почвах (литосолях) и фрагментами нивального пояса (небольшие ледники, фирновые поля).
Глава III. Региональные различия внутриландшафтной дифференциации беспозвоночных экотона высокогорий
Первоначально мы хотели бы остановиться на региональных особенностях выбранных для исследованиях участках – хребтах Малая Хатипара и Абишира-Ахуба и долине реки Птыш.
Хребет М. Хатипара является северным отрогом Главного Кавказского хребта и образует в пределах Тебердинского заповедника часть левого склона долины реки Теберды. Хребет представляет собой северное крыло антиклинали, замковая часть которой размыта рекой Большая Хатипара.
Современный рельеф хребта создавался под влиянием разнообразных факторов — геологического строения, древнего оледенения, водной и лавинной эрозии, и имеет морфологическую зональность. Расположение морфологических зон совпадет с положением древних снеговых границ (Тушинский, 1957). Нижняя треть склонов хребта представляет собой участки троговой долины Теберды. В нижней части они круты и сложны пролювиально-коллювиальными и флювиогляциальными отложениями. На отметках 2300 м имеются переломы склонов (плечи трогов), переходящие в относительно пологие участки с распространением древних цирков и каров. Затем снова следуют более крутые поверхности, переходящие на высотах 2600-2700 м в плоские площадки каров.
Большую роль в формировании рельефа играла водная лавинная эрозия. Восточный склон хребта расчленен глубокими речными долинами Малой и Большой Хатипары, которые являются левыми притоками р. Теберды. В целом, в пределах хребта, преобладающими формами рельефа являются крутые склоны вторичных долин и балок, сложенные коллювием, конуса выноса и ледниковые формы (кары, морены).
Значительные колебания относительных высот в пределах хребта обусловили формирование вертикальных различий климата, растительности, почв и животного мира (основные показатели рассмотрены во второй главе).
Изменение с высотой климатических условий определяет дифференциацию растительного и почвенного покрова и, как следствие, формирование высотных геоботанических поясов. На склонах древних трогов р. Теберды и долин второго порядка произрастают пихтово-сосновые леса на бурых горно-лесных почвах. В нижней трети склонов трогов и на днище долин получает распространение растительность переходного облика. Здесь совместно с хвойными (пихтово-сосновыми) произрастают буковые и буково-дубовые леса. В верхней трети склонов трогов, выше пояса хвойных лесов, растут парковые сосновые редколесья па грубо скелетных бурых горно-лесных почвах, березовые криволесья на горно-кустарниковых почвах и фрагменты субальпийских лугов на горно-луговых почвах. С a6cолютных высот 2300 м начинаются луговые ассоциации: субальпийские и альпийские луга на горно-луговых почвах. Вершина хребта занята субнивальным поясом. (Шальнев, 1973).
Хребет Абишира-Ахуба представляет собой глубокорасчлененные крутосклонные, эрозионно - тектонические горы Передового хребта, с реликтами древнего оледенения, с субальпийскими и альпийскими лугами на бурых горно-луговых почвах, со скалами, осыпями и каровыми озерами (Савельева, 1973).
Долина реки Птыш в пределах северного склона Водораздельного хребта Большого Кавказа по рельефу является составной частью Южно-Юрской депрессии (Домбайский участок) с абсолютными отметками 1800-1900м над уровнем моря. Это троговая долина, у которой склоны сложены метаморфическими породами палеозоя, а днище заполнено четвертичными моренными, флювиогляциальными и аллювиальными отложениями. Склоны трогов при переходе к днищу выположены конусами выноса эрозионных ложбин, сложенных коллювиально-пролювиальными отложениями. На правом борту долины эти конуса выноса формируют общий шлейф, а на левом борту выражены слабо, так как размываются рекой Птыш.
В долине Птыша хорошо сохранились следы последней стадии четвертичного оледенения, получившего название 3-его этапа каракельского оледенения, его 2-ой фазы (Каракельской) и Аманаузской стадии (Тушинский, 1949, Сафронов, 1960). Длина ледника Аманаузской стадии достигла 5,0-5,5км (Сафронов, 1960). У входа в долину сохранилась мощная конечная морена (высотой до 40м) последней стадии оледенения. Далее на днище долины сохранились еще три невысоких конечных морены (3-5м высотой), которые фиксируют стадии отступления ледника постаманаузской стадии. Хорошо выражены донные и боковые морены современной стадии Фернау отступания ледника.
Река Птыш берет начало с Птышского ледника северного склона Водораздельного хребта. Профиль реки пологий, слабо врезанный в днище, поэтому в рельефе представлены только фрагменты пойменных верхнечетвертичных террас. В нижней части течения (конечная морена Аманаузского оледенения) сформировалась «четка» с широкой поймой на месте бывшего озера.
В связи с абсолютными высотами долины и климатическими условиями в долине сформировалась растительность переходного геоботанического пояса от хвойных (пихтовых) лесов к субальпийским лугам (Шальнев, 2005). Здесь произрастают березовые криволесья на конечной морене (Аманаузская стадия) и частично на склонах трога. Большая часть днища долины занята субальпийскими лугами (Шальнев, Конева, 2006).
Ландшафты же хребта относятся к окультуренным природным (Шальнев, 2004). Такие ландшафты чрезвычайно важны для природы и человека и требуют подробного изучения, поскольку в них проявляются естественные природные закономерности.
3.1. Беспозвоночные Тебердинского биосферного государственного заповедника
Тебердинский биосферный государственный заповедник был выбран в качестве исследуемой нами территории, в основном из-за высокой степени сохранности естественных экосистем, которые обусловливают высокое видовое разнообразие животного мира Западного Кавказа. Из всех исследуемых нами ключевых участков хребет Малая Хатипара является эталонным участком не испытывающим антропогенного прессинга (Шкарлет, Разумов, 2003).
Учет численности беспозвоночных - обитателей травяного покрова проводился методом кошения энтомологическим сачком. Данный метод был выбран для получения данных о соотношении обитателей травостоя и изменениях численности беспозвоночных в травостое в течение определенного времени (Программа …, 1974). Подсчет беспозвоночных проводился на 100 взмахов сачком. Для обеспечения большой точности были выбраны следующие параметры сачка: длина ручки - 1 м, диаметр обруча - 30 см, длина мешка - 70 см. Кошение проводилось после схода росы с 11 часов по 15 часов в период наибольшей активности населения травостоя. Оптимальное количество взмахов подбиралось опытным путем из предлагаемых различными авторами методик (Фасулати, 1971; Чернов, Руденская 1970; Второв, 1968).
Определение проходило в условиях камеральной обработки. Нами беспозвоночные были определены до уровня семейств посредством определителей: «Определитель насекомых европейской части СССР», в пяти томах, под ред. Г.Я. Бей-Биенко; Мамаев Б.М. «Определитель насекомых по личинкам»; Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. «Определитель насекомых Европейской части СССР» (1970); Горностаев Г.Н. «Насекомые СССР» (1970); http://flora-world.by.ru/; http://www.floranimal.ru/orders/2712/; http://zoo. Помощь в определении видовой принадлежности собранного материала оказывал с.н.с. ЮНЦ РАН, к.б.н. Ю.Г. Арзанов.
Исследования проводились на трех ключевых участках, расположенных в пределах Бокового и Передового хребтов Западного Кавказа, где были выделены следующие основные виды контурных экотонов: водораздельный, склоновый и ледниковый (Шальнев, Нефедова, 2006).
3.1.1. Беспозвоночные хребта Малая Хатипара
Водораздельный экотон речных долин 2-го порядка (притоков Теберды) характеризуется линейным простиранием, имеет четко выраженные склоны и гребень (рис. 4). Данный вид экотона был рассмотрен на примере хребта Малая Хатипара. Пояс экотона представляет переходную ленточную границу от ландшафтов хвойных лесов троговой долины реки Теберды к высокогорным ландшафтам лугов. До 2200 м. над уровнем моря на склонах долин юго-восточной экспозиции распространены сосновые леса, переходящие выше в сосновые редколесья. Пихтовые леса северной экспозиции поднимаются до 2000 м. над у.м. и замещаются березой. Выше расположен пояс субальпийских лугов.
Изучение внутриландшафтной дифференциации насекомых проводилось на примере анализа таксономических групп насекомых хортобионтов в ранге семейств, с выделением доминантных видов или родов. Анализ населения насекомых проведенный по этим таксонам позволяет выявить основные особенности структуры населения насекомых выделенных видов экотона. Особое значение такие исследования приобретают в условиях высокогорных ландшафтов, которые при современном уровне антропогенной трансформации природных комплексов играют важнейшую роль в сохранении генофонда биологического разнообразия и выступают в качестве эталонных участков биосферы (Конева, 2007).

Рис. 4. Фрагмент ландшафтной карты водораздельного экотона
хребта Малая Хатипара.
Легенда:Урочища:
22 – верхней части очень крутых склонов юго-западной экспозиции с сосновыми редколесьями и полянами высокотравных лугов на примитивных бурых горно-лесных и горно-луговых почвах;
23 – верхней части очень крутых склонов северо-восточной экспозиции с березовыми криволесьями и зарослями рододендрона на бурых дерновых горно-лесных и горно-кустарниковых почвах;
30 – верхней части склонов юго-западной экспозиции с разнотравно-злаковыми лугами на дерново-луговых субальпийских почвах;
31 – крутых склонов балок третьего порядка северо-восточной экспозиции с разнотравно-злаковыми лугами и зарослями рододендрона на горно-луговых и горно-кустарниковых почвах.
Первоначально мы рассмотрели структуру отрядов хортобионтов, присущих водораздельному экотону хребта Малая Хатипара (рис. 5)
Можно увидеть, что численно преобладает отряд Homoptera – 32% от общего числа насекомых отрядов. Хотя перевес над количеством других отрядов наблюдается незначительный – отряд Hemiptera (29%) и отряд Coleoptera (27%). Более детальное отличие мы отобразили на последующих круговых диаграммах, которые показывают структуру семейств относительно каждого отряда. Причем подчеркнутым шрифтом отмечены фоновые виды или рода для каждого семейства. Минимальное процентное соотношение (12%).

Рис. 5. Структура отрядов хортобионтов для водораздельного экотона хребта Малая Хатипара.
Наибольшим видовым разнообразием Coleoptera выделяется именно водораздельный экотон, по сравнению с ледниковым и склоновым видами экотона (рис. 6).

Рис. 6. Структура населения Coleoptera для водораздельного экотона хребта Малая Хатипара.
Отряд Coleoptera отличается наибольшим видовым разнообразием, хотя и не большей численностью среди остальных отрядов данного вида экотона. Это объясняется тем, что это наиболее «заповедный» участок Тебердинского заповедника, почти неизмененный человеческой деятельностью. Доминантным семейством из отряда Coleoptera является семейство Curculionidae с основными видами и родами Authonomus rubi, Nastus albidusovoideus, Apion vicial, Zacladus geranii, Nastus fausti, Otiorhynchus tatarchani, Otiorhynchus kirschi, Alans sp. Данная группа составляет значительную часть хортобионтов. Многочисленными являются семейство Staphylinidae (Anthophagus angusticollis roubali и Eusphalerum sp.) и семейство Elateridae (Athous sp., Dascillus cervinus). Выделяются и семейство Oedemeridae (Oedemera lateralis, Oedemera flavescens, Chrysanthia viridisima), семейство Coccinellidae (Coccinula quatuordecimpunctata, Coccinella septempunctata, Vibidia duodecipunctata, Subcoccinella vigintiquatuorpunctata), семейство Cantharidae (Rhagonycha limbata, Rhagonycha translucida), семейство Cerambycidae (Anastragalia dubia, Phytoecia affinisboeberi, Agapanthia villosoviridescens, Agapanthia intermedia, Stenurella jaegeri, Brachyta interrogationis), семейство Chrysomelidae (Cryptocephalus sericeus, Cryptocephalus ctibratus, Bromius obscurus). Семейство Листоедов здесь весьма малочисленно, но они присутствуют. А так как насекомые из семейства Chrysomelidae служат своеобразными индикаторами антропофитной растительности, в частности, это конский щавель, чемерица Лоббиля. Значит, М. Хатипара в недалеком прошлом подвергалась значительной антропогенной нагрузке.
Нам удалось также проанализировать и структуру насекомых Hemiptera (рис.7).

Значительно преобладает в отряде Hemiptera семейство Miridae с доминантным родом Polimerus sp. Семейство Pentatomidae составляет практически четверть от общего состава Hemiptera и представлено видами – Dolicoris baccarum, Peribalus vernalus, Carpocoris melanocerus. Семейство Reduviidae представлено одним видом – Rhynocoris iracundus и семейство Scutelleridae также отмечено наличием только одного вида – Eurygaster maura.
Далее хотелось бы остановиться на преобладающем по численности отряде – Homoptera, к сожалению, определение было осуществлено только до уровня семейств (рис. 8).
Более половины беспозвоночных отряда Homoptera представлены семейством Cicadellidae sp. Также выделяется семейство Psyllidae sp. и встречаются беспозвоночные семейства Cercopidae (Cercopis sanguinea) представители подотряда Aphidinea.

Рис. 8. Структура населения Homoptera для водораздельного экотона хребта Малая Хатипара.
Отряд Orthoptera явился самым малочисленным из рассмотренных выше отрядов. Но в отличие от всех остальных видов экотона, самым «полноценным», т.е. представленным наличием двух ведущих семейств прямокрылых – семейства Tettigoniidae и семейства Acrididae, причем особи отмечены уже во взрослом состоянии (рис. 9).

Рис. 9. Структура населения Orthoptera для водораздельного экотона хребта Малая Хатипара.
Отряд Orthoptera включает достаточно богатый состав видов и доминирует семейство Tettigonidae с основыми видвми - Poecilimon heroicus, Euconocercus caucasicus, Pholidoptera fallax, который является эндемиком Кавказа и Decticus verriuccivorus. Семейство Acrididae отмечено двумя видами – Chrysochraon dispar и Aeropedellus variegates (рис.9).
В прошлом на территории заповедника велась весьма активная антропогенная деятельность, связанной с выпасом скота, о чем свидетельствуют достаточно обширные площади, занятые антропофитами и высокотравьем – 55% в нижней части и 15% в верхней части экотонного участка (рис. 10). Если рассматривать распространение антропофитов по высотному профилю от нижней границы экотона к верхней, то антропофитная растительность является следствием существования в днище цирка когда-то коша. На верхней границе экотона антропофиты вытесняются типичными субальпийскими видами. Это говорит о восстановлении биоценоза за время существования заповедника (Олейникова, 2005).

Рис. 10. Соотношение антропофитной растительности в пределах границ водораздельного экотона хребта Малая Хатипара.
Всего за период исследования на хребте Малая Хатипара было отловлено 409 особей. Из рассматриваемых отрядов, представленных на ключевом участке водораздельного экотона (Coleoptera, Orphoptera, Hemiptera и Homoptera), наиболее богатым видами является отряд Coleoptera и отряд Orthoptera. Отряды Hemiptera и Homoptera выделяются численным преобладанием – 29% и 32%, соответственно. Наибольшее разнообразие хортобионтов все же исследовано на территории водораздельного экотона хребта Малая Хатипара, где произрастают различного рода сосновые, пихтовые, смешанные хвойно-широколиственные и лиственные леса с травяным покровом разной мощности, что является существенным фактором местообитания изучаемой нами группы беспозвоночных (Конева, 2005).
3.1.2. Ландшафтно-экологический подход в изучении беспозвоночных на уровне фаций (на примере водораздельного экотона хребта Малая Хатипара)
Ландшафтно-экологический подход актуален в изучении беспозвоночных. Рассматривались насекомые водораздельного экотона на уровне фаций луговой растительности различных экспозиций (обоснование дано в главе II). Ведущим фактором явились микроклиматические показатели, полученные В.А. Шальневым на стационаре Тебердинского заповедника в 1971 г. Были выбраны три фации луговых ассоциаций различных экспозиций: северной, восточной и южной (рис. 11).
Рис. 11. Ландшафтная карта фаций Тебердинского ландшафта
(Шальнев, 1971).
Легенда:
- Урочище верхней трети крутых склонов долины М. Хатипары северо-северо-восточной экспозиции, сложенное коллювием, состоит из фации:
1) разнотравно-пестрокостровые луга на горно-луговых мощных сильно дерновых скелетных почвах;
- Урочище верхней трети крутых склонов долины М. Хатипары северо-северо-восточной экспозиции, сложенное коллювием и выделяется фация:
2) разнотравно-злаковые луга на горно-луговых мощных сильно дерновых скелетных почвах;
- Урочище древнего цирка восточной экспозиции, сложенное коллювием:
3) верхняя часть склона с вейниково-злаково-разнотравной растительностью на горно-луговых мощных сильно дерновых скелетных почвах.
Луг северный занимает пригребневые участки склона северо-северо-восточной экспозиции, крутизной 16-18º, с абсолютной высотой 2350 м. Растительность – разнотравно-злаковые луга. В почвенном покрове горно-луговые дерновые скелетные почвы. Вторая фация (луг восточный) располагается в верхней трети склона древнего цирка на склоне восточной экспозиции, крутизной 20º. Растительный покров имеет смешанный видовой состав, характерный для склонов и северных, и южных экспозиций. Растительную ассоциацию можно назвать вейниково-злаково-разнотравной. В почвенном покрове горно-луговые мощные скелетные почвы. Третья фация (луг южный) занимает пригребневой участок склона юго-юго-восточной экспозиции, крутизной 31º, с абсолютной высотой 2380 м. В почвенном покрове развиты горно-луговые мощные сильнодернованные почвы. Растительность представлена разнотравно-пестрокостровым лугом.
Ведущим фактором явились микроклиматические показатели, на которые в свою очередь, влияет экспозиция склонов. Она в первую очередь влияет на дневной и суточный ход составляющих радиационного и теплового балансов. Максимальные показатели радиационного баланса приходятся на луг южный в полдень – 1,2 кал/см2. На лугу восточном и северном он быстро растет в утренние часы и своего максимума достигает уже к десяти часам – 1,1 кал/см2 и 0,6 кал/см2, соответственно. Если же рассматривать показатель затрат тепла на испарение, то максимум тепла на лугу южном тратится на околополуденные – 0,55 кал/см2, на лугу северном и восточном – в утренние часы – 0,45 кал/см2 и 0,58 кал/см2. Еще более показательны суммы дневных и месячных величин. Хорошо обеспечен теплом луг южный. Приход суммарной радиации за ясный день здесь составляет 777 кал/см2, а за месяц 16,6 ккал/см2. Северный луг получает суммарной радиации в ясный день только 519,5 кал/см2, что на 33% меньше луга южного. Месячная ее величина равняется 11,3 ккал/см2. Луг восточный обеспечен теплом лучше луга северного, так как приход суммарной радиации за день составляет 662,4 кал/см2. При сравнении суточных величин составляющих теплового баланса эти закономерности сохраняются. Самые большие затраты тепла на испарение (271,0 кал/см2), поток в почву (10,0 кал/см2) и турбулентный поток в воздух (214,6 кал/см2) отмечаются на лугу южном. Соотношение же составляющих частей радиационного баланса по составляющим теплового баланса в каждой фации разное. Если на лугу южном затраты тепла на испарение составляют 54% от общей величины радиационного баланса, а на турбулентный поток в воздух расходуется только 43%, то на лугу северном затраты тепла на испарение растут до 64% и составляют 171,2 кал/см2, а на турбулентный поток в воздух они снижаются до 35% и составляют 89,9 кал/см2. На лугу восточном эти соотношения почти одинаковы и составляют – 185,6 кал/см2 и 187,6 кал/см2 в сутки, соответственно (Шальнев, 1971).
Особенности радиационного и теплового баланса влияю на термический и водный режим фаций. Лучше всего градиентные различия температур воздуха проявляются на лугу южном. Если на высоте 2 м температура воздуха в полдень ясного дня поднимается до 14º, то в приземном слое (0,2 м) она возрастает до 17,8º, а на поверхности деятельного слоя – до 20,6º.На склоне северной экспозиции градиентные различия температур не так велики. На высоте 2 м воздух в полдень прогревается только до 11,5º, а на уровне 0,2 м до 13,6º. Хорошо видны градиентные различия в разных фациях по средним суточным величинам: и температуре воздуха, и абсолютной влажности. Исходя, из особенностей хода средних суточных величин температуры воздуха и почвы и абсолютной влажности воздуха на разных высотных уровнях в ясные дни и расположение на склонах различной экспозиции следует, что лучше всего прогревается слой почвы на глубине 0-10 см. Здесь отмечаются самые высокие средние суточные температуры. На лугу южном на глубине 5 см они равняются 12,8º. В то же время температура воздуха на высоте 2 м составляет 9,1º. Наименьшие колебания температур в почвенном слое наблюдаются на склонах восточной экспозиции: на глубине 0,10 см – 12,2º, а на высоте 2 м – 8,8º.
Термические условия почв влияют на режим влажности, поэтому почвы луга южного являются самыми «сухими». Так, на глубине10 см содержание влаги составляет 40%. На лугу северном ее содержание возрастает до 60%. Высокий процент влажности в почвах обусловлен большим количеством атмосферных осадков – 108 мм. Обычно процент влажности в горизонте А бывает ниже, например, на лугу южном влажность составляла 25%, а на северном – 42%.
Ветровой режим фаций в горах во многом зависит от экспозиции склона. В целом, наблюдался юго-западный вынос воздушных масс. На склонах южной экспозиции он усиливался восходящим потоком горно-долинных ветров, поэтому на лугу южном отмечались самые высокие скорости ветра. Средние суточные величины за ясные дни на высоте 2 м составляли 2,9 м/сек., а на высоте 0,5 м – 1,2 м/с. Иногда скорости достигали 11 м/с. Повышения скорости ветра на склонах южной экспозиции наблюдались в летние месяцы. На склонах северной экспозиции уменьшаются не только скорости ветра, но и меняется его направление. Так, на лугу северном преобладали ветры северных направлений, и средние суточные величины их в ясные дни были не больше 1,6 м/сек. Максимальные скорости не превышали 3,2 м/сек. На лугу восточном средние суточные скорости выше и равняются 2,6 м/сек.
Режим ветра оказывает большое влияние на термические условия приземного слоя воздуха. На лугу южном значительные скорости ветра в дневные часы приводят к усилению турбулентного обмена и выносу тепла за пределы фации. Поэтому средние суточные величины температур воздуха на высоте 2 м лишь на 1-2º выше, чем на лугу северном. Передача тепла турбулентным потоком в воздух достигает здесь больших величин – 43% общей суммы радиационного баланса. Не последнюю роль ветровой режим играет и в снижении процента влажности почв южного луга.
Растительный покров описываемых фаций относится к субальпийским лугам. Территория луга южного занята разнотравно-пестрокостровой ассоциацией. Здесь наиболее характерными представителями являются: костер пестрый, овсяница пестрая, сон-трава фиолетовая, подорожник, лапчатка, пуповка Рудольфа и др. Всего произрастает 74 вида растений. Для этого луга к растительным ассоциациям «привязаны» следующие виды хортобионтов: отряд Orthoptera – 23 особи от общего количества, из которых семейству Tettigoniidae относится вид Poecilimon heroicus, а к семейству Tettigoniidae – вид Chrysochraon dispor. Отряд Homopteta – 15 особей семейства Cercopidae c одним видом Cercopis sanguinea. Отряд Hemiptera – 22 особи cемейства Pentatomidae с доминантными видами - Dolicoris baccarum, Peribalus verualis, Carpocoris melanocerus. К отряду Coleoptera относится 9 насекомых, из которых семейству Curculionidae принадлежат два вида - Zacladus geranii и Apion vicial, семейству Cerambycidae – виды Phytoecia (Mussaria) affinisbolberi и Anastrangalia dubia, семейству Oedemeridae – вид Oedemera flavescens (рис. 12).

Рис. 12. Распределение отрядов и доминантных видов хортобионтов на лугу южной экспозиции.
На лугу северном произрастают разнотравно-злаковые ассоциации с преобладанием вейника тростниковидного, мятлика длиннолистного, ветреницы пучковатой, купальницы и др. Все вышеперечисленные факторы способствуют распространению здесь таких же семейств, что и для южного луга, но с некоторыми отличиями, также отличается и набор видов. Всего 62 вида растений.
Основными доминатными отрядами по количественным характеристикам является отряд Homopteta – 41 особь, причем более половины всех насекомых этого отряда находятся на стадии личиночного развития и отряд Coleoptera, который к тому же имеет и достаточно богатый видовой набор. Он представлен 25 особями, которые относятся к двум семействам: к семейству Curculionidae принадлежат два вида - Zacladus geranii и Nastus fausti, а к семейству Oedemeridae – вид Chrysanthia viridissima. Отряд Orthoptera представлен в количестве 21 особи, которые все находятся на личиночной стадии развития и видовую принадлежность трудно идентифицировать. На северном лугу малочисленным отрядом является отряд Hemiptera – 13 особей cемейства Pentatomidae с доминантным видом - Dolicoris baccarum (рис. 13).

Рис. 13. Распределение отрядов и доминантных видов хортобионтов на лугу северной экспозиции.
На лугу восточном распространены злаково-разнотравные луга, число видов которых превышает 80. Здесь отмечена 81 особь хортобионтов. Доминируют два отряда – отряд Homopteta и отряд Hemiptera по 27 особей. Как и на предыдущих участках преобладающими по численности являются именно эти два отряда, большая часть которых представлена насекомыми, находящимися на личиночной стадии развитии. В отряде Hemiptera также выделяется ранее не встречающийся вид Authonomus rubi и род Polimerus sp. Отряд Coleoptera (18 особей) и здесь выделяется по видовому разнообразию и представлен четырьмя семействами - семейством Oedemeridae – вид Chrysanthia viridissima, семейство Coccinellidae – вид Subcoccinella vigintiquatuorpunctata, семейство Elateridae – род Athous и семейство Curculionidae – вид Authonomus rubi. И самым малочисленным отрядом и на восточном лугу явился отряд Orthoptera, большинство особей представлено личинками и только две взрослые особи вида Chrysochraon dispor (рис. 14).

Рис. 14. Распределение отрядов и доминантных видов хортобионтов на лугу восточной экспозиции.
В итоге мы предоставляем выборку общих видов для трех рассмотренных экспозиций (рис.15).

Рис. 15. Соотношение видов насекомых на южном, северном и восточном склонах.
При общем анализе данных по сравнению насекомых, выявлено количество общих видов для всех фаций водораздельного экотона только для семейства Cercopidae – вид Cercopis sanguinea. Общими для северной и южной экспозиций является вид Dolicoris baccarum cемейства Pentatomidae и вид Zacladus geranii семейства Curculionidae. Вид Carpocoris melanocerus cемейства Pentatomidae и вид Chrysochraon dispor семейства Acrididae характерны для южной и восточной экспозиций. А на склонах северной и восточной экспозиций встречаются насекомые отряда Homopteta и отряда Orthoptera, но только на личиночной стадии развития. Все выше перечисленные виды встречаются только на одной определенной фации и больше не дублируются. Такое закономерное распределение видового набора хортобионтов доказывает прямую взаимосвязь микроклимата, растительных ассоциаций и хортобионтов.
3.1.2. Беспозвоночные долины реки Птыш
Ледниковый экотон рассматривался на примере троговой долины реки Птыш. Склоны трогов при переходе к днищу выположены конусами выноса эрозионных ложбин. В долине Птыша хорошо сохранились следы последней стадии четвертичного оледенения, получившего название 3-его этапа каракельского оледенения (Тушинский, 1949, Сафронов, 1960). У входа в долину сохранилась мощная конечная морена (высотой до 40м.) последней стадии оледенения. Далее на днище долины сохранились еще три невысоких конечных морены (3-5м высотой), которые фиксируют стадии отступления ледника постаманаузской стадии. Хорошо выражены донные и боковые морены современной стадии «фернау» отступания ледника (Шальнев, Конева, 2006).
В связи с абсолютными высотами долины и климатическими условиями в долине сформировалась растительность переходного геоботанического пояса от хвойных (пихтовых) лесов к субальпийским лугам (Шальнев, 2005). Здесь произрастают березовые криволесья на конечной морене (Аманаузская стадия) и частично на склонах трога. Большая часть днища долины занята субальпийскими лугами (рис. 16).
![]()

Рис. 16. Фрагмент ландшафтной карты ледникового экотона
долины реки Птыш.
Легенда:
Урочища:
А. Сложное урочище днища долины, сложенное аллювием и флювиогляциальными отложениями. Состоит из простых урочищ:
3 – конечная морена, частично размытая рекой (левый борт), с березовой растительностью на примитивных грубоскелетных почвах;
4 - конечные морены; частично перекрытые осыпями, с разнотравно-злаковой растительностью на примитивных горно-луговых почвах;
Б. Сложные урочища склонов троговой долины западной и восточной экспозиции, сложенные коллювиально-пролювиальными отложениями и кореннымит породами (гнейсами). Состоит из простых урочищ:
6 - конуса выноса, сложенные коллювиально-пролювиальными отложениями, западной (восточной) экспозиции с субальпийскими лугами и березовыми криволесьями на горно-кустарниковых почвах;
7 - конуса выноса, сложенные коллювиально-пролювиальными отложениями, западной экспозиции с луговой субальпийской растительностью на горно-луговых почвах;
8 – то же, восточной экспозиции с луговой субальпийской растительностью и пятнами рододендрона.
Была рассмотрена структура отрядов хортобионтов, присущих ледниковому экотону долины реки Птыш (рис. 16).
Отметим, что численно преобладает отряд Homoptera – 56% от общего числа насекомых отрядов. Причем более 70% насекомых отряда находится на личиночной стадии развития. Далее следует отряд Hemiptera (38%). Представителей последнего отряда в 1,5 раза меньше, чем равнокрылых. Для отряда также характерно доминирование особей личиночной стадии развития – более 70%. Отмечено совсем малое количество насекомых, относящихся к отряду Coleoptera. Их всего 6% от всех отловленных особей. Самым малочисленным оказался отряд Orthoptera – менее 1%, представленными личинками. Более детальное отличие мы отобразили на последующих круговых диаграммах, которые показывают структуру семейств относительно каждого отряда. Причем подчеркнутым шрифтом отмечены фоновые виды или рода для каждого семейства (рис. 17).

Рис. 17. Структура отрядов хортобионтов для ледникового экотона долины реки Птыш.
Отряд, Coleoptera занимающий предпоследнее место (6%), характеризуется достаточно большим видовым разнообразием, хотя и гораздо меньшим, чем на водораздельном экотоне. Отряд Сoleoptera представлен пятью семействами (рис. 18).
Доминантным семейством является семейство Cantharidae с видами Cantharis sudetica, Rhagonycha limbata и. Для семейства Curculionidae – Phyllobius argeutatus, Otiorhynchus tatarchani, Otiorhynchus bideutatus, Zacladus geranii. Менее распространенными явились семейство Chrysomelidae с видами и родом Gastroidea viridula, Cryptocephalus sp. И семейство Coccinellidae с одним отмеченным здесь видом Coccinula quatuordecimpunctata.

Рис. 18. Структура населения семейств Coleoptera для ледникового экотона долины реки Птыш.
Нам удалось также проанализировать и структуру насекомых Hemiptera (рис. 19).

Рис. 19. Структура населения семейств Hemiptera для ледникового экотона долины реки Птыш.
Отряд Hemiptera представлен только одним семейством Miridae и единственным здесь родом Polimerus sp. Для наглядного сравнения нами был выбран именно этот тип диаграммы потому, что можно отметить, что более 70% особей находится на стадии личиночного развития.
Далее нами был рассмотрен преобладающий отряд Homoptera (рис. 20).

Рис. 20. Структура населения семейств Homoptera для ледникового экотона долины реки Птыш.
Единственным выделенным здесь семейством является семейство Cicadellidae. К сожалению, невозможно было определить даже род особей, но более 70% беспозвоночных являются взрослыми особями.
На рисунке 21 представлена структура отряда прямокрылых. Группа Orthoptera также малочисленна и представлена, в большей степени особями, которые находятся на личиночной стадии развития. Но все же нами было отмечено семейство Pamphagidae и определен вид – Nocaracris cyanipes или каменная кобылка, встречающаяся на альпийских лугах и скалистых участках с обилием мхов и лишайников. Местообитания каменной кобылки (Nocaracrus cyanipes) приурочены к альпийским коврам и каменистым пустошам у границ ледников на высотах 2800 – 3200 м. Следствием адаптаций прямокрылых к штормовым ветрам высокогорий является наличие бескрылых видов и форм с частично редуцированными крыльями.

Рис. 21. Структура населения семейств Orthoptera для ледникового экотона долины реки Птыш.
В принципе, существует вполне строгая закономерность дифференциации насекомых травяного покрова. По мере постепенного отступания ледника, происходит последовательное заселение освобожденной территории. Сначала появляется березы, как более устойчивая к неблагоприятным условиям порода, позже антропофитная растительность, а потом происходит распространение и насекомых, в прямой зависимости от растительных ассоциаций. Однако территориальные различия травянистой растительности по долине реки Птыш различны, что связано с временными процессами зарастания моренных отложений. В то же время, травянистая растительность данного ледникового экотона, имеет значительную схожесть по видовому составу с субальпийскими лугами на конусах выноса долины как донора видового разнообразия. Отсюда и зависимость в распространении беспозвоночных как ледниковых представителей - вид Nocaracris cyanipes, так и субальпийских – вид Coccinula quatuordecimpunctata или вид Zacladus geranii.
При характеристике антропогенной нарушенности, можно отметить, что она здесь отсутствует. Хотя представители антропофитов отмечены на донной морене долины реки и на конусах выноса, расположенных в непосредственной близости от самого ледника (рис. 22). Присутствие нтропофитной растительности объясняется их способностью к заселению территории после освобождения морен после схода ледника.

Рис. 22. Соотношение антропофитной растительности и разнотрано-злаковой в пределах границ ледникового экотонадолины реки Птыш.
Все же на 75% преобладает разнотравно-злаковая растительность, а антропофиты представлены 25 %, причем фоновым растением для донной морены является борщевик, а для конусов выноса – чемерица Лоббиля и чертополох. Также индикаторами антропофитности являются и беспозвоночные – личинки и взрослые особи Gastroidea viridula семейства Chrysomelidae.
В целом, многочисленность насекомых (отряды Homoptera, Hemiptera) с преобладанием доли личинок, уравновешивается бедностью видового разнообразия и малочисленностью отряда Coleoptera и присутствием Orthoptera (на стадии личиночного развития) с типичным ледниковым видом – Nocaracris cyanipes.
3.2. Беспозвоночные хребта Абишира-Ахуба
Склоновый экотон рассмотрен на примере южного склона хребта Абишира-Ахуба слабой эрозионной расчлененности. Он является одним из наиболее выраженных видов ленточного типа экотона, который находится на границе двух доминантных геоботанических поясов – сосновых лесов и субальпийских лугов (рис. 23). Сосновые леса среднегорий Архызской котловины распространены до отметки 2200 м. над уровнем моря. Выше они переходят в небольшие массивы сосновых редколесий, разделенных субальпийскими лугами, которые приурочены в большей части к древним карам, заполненным коллювием. От них вниз по склону протягиваются эрозионные ложбины, разделяющие сосновые леса на отдельные массивы. Выше 2300-2350 м. над уровнем моря начинается пояс субальпийских лугов.
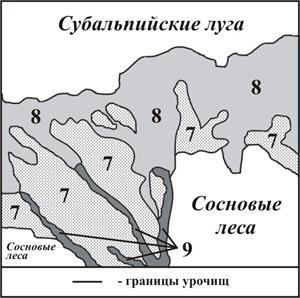
Рис. 23. Фрагмент ландшафтной схемы склонового экотона
хребта Абишира-Ахуба (южный склон).
Легенда:
Урочища:
7 – очень крутых склонов с обнажением коренных пород и коллювиальными осыпями, с сосновыми редколесьями на грубоскелетных примитивных бурых горно-лесных почвах;
8 – древних каров, сложенных коллювием, с субальпийскими лугами на горно-луговых почвах;
9 – эрозионных и лавинных ложбин, сложенных коллювиально-пролювиальными отложениями с луговой растительностью.
Была рассмотрена структура отрядов хортобионтов, присущих склоновому экотону хребта Абишира-Ахуба (рис. 24). Доминирует отряд Homoptera – 60%, как на ледниковом экотоне. Перевес этого отряда над следующим – отрядом Hemiptera составляет более 30%. Насекомые отряда Coleoptera составляют 11% от общего состава хортобионотов хребта Абишира-Ахуба. Самым малочисленным, как и на всех представленных видах экотона, явился отряд – Orthoptera – всего 3% от всех беспозвоночных.
Такая структура соотношения отрядов вполне оправдана для территории, ввиду антропогенного фактора.

Рис. 24. Структура отрядов хортобионтов для склонового экотона хребта
Абишира-Ахуба.
Отряд Coleoptera представляет собой довольно интересное сочетание семейств и набор видов, не отмеченный в зоне экотона хребта Малая Хатипара и в долины реки Птыш (рис. 25). Примечательно, что наиболее разнообразно сем. Curculionidae в экотонной зоне хребта Абишира-Ахуба при наименьшей площади исследованной нами территории. Поэтому доминирует семейство Curculionidae – 38%, с основными видами и родами – вид Cionis hartulanus доминанта, также отмечены и виды - Nastus fausti, Zacladus geranii, Otiorhynchus psudobrachyalis. 20% отловленных особей приходится на семейство Chrysomelidae – 20%. Особенностью представленного комплекса является не полное доминирование семейства листоедов среди остальных семейств, но оно представлено на втором месте и составляет 20% от общего числа. Хотя видовая структура все же не однообразна – это личинки и взрослые особи вида Gastroidea viridula и насекомые рода Cryptocephalus (его виды – Cryptocephalus moraei, Cryptocephalus bipunctatus). Такая структура населения склонового экотона – результат сильной пастбищной нагрузки. В долине идет летнесезонный выпас как крупного рогатого скота, так и мелкого – овец, коз. Естественные сообщества долины заменены антропогенными. Растительность днища представлена антропофитами – щавелем конским, крапивой двудомной и др. Соотношение семейства Elateridae (Dascillus cervinus, Athous sp.) и семейства Staphylinidae (Anthophagus angusticollis roubali и Eusphalerum sp.) составляет по 15 %, причем в отличие от хребта Малая Хатипара, здесь доминирует в семействе Elateridae вид Dascillus cervinus, а не род Athous sp. Видовой состав семейства Staphylinidae одинаков. Причем ни семейства щелкунов, ни семейства стафилинов не отмечено в долине реки Птыш. Также для склонового экотона отмечены и семейства – Mordellidae, Oedemeridae (Oedemera flavescens) и выявлено только здесь семейство Alleculidae (вид Hymenalia rufipes).

Рис.25. Структура населения семейств Coleoptera для склонового экотона хребта Абишира-Ахуба.
Также нами проанализирована структура отряда Hemiptera для экотонной зоны хребта Абишира-Ахуба (рис. 26). Более 90% всех особей отряда полужесткокрылых составляет семейство Miridae с доминантным родом Polimerus sp.), но только здесь встречается еще один вид Adelphocoris lineolafus. Только на хребте Абишира-Ахуба отмечено семейство Coreidae c единственным видом Syromastus rhombeus. Выделены и насекомые семейства Pentatomidae, которое представлено видом – Dolicoris baccarum.

Рис. 26. Структура населения семейств Hemiptera для склонового экотона хребта Абишира-Ахуба.
При рассмотрении отряда Homoptera (рис. 27) выявлено, что практически одинаковое соотношение двух семейств в отряде. Незначительно доминирует семейство Cicadellidae sp. – 55%. 45% насекомых отряда равнокрылых представлены, как и в остальных регионах, семейством Psyllidae sp. В отличие от остальных видов экотона на хребте Абишира-Ахуба не выявлено семейство Cicadellidae.

Рис. 27. Структура населения семейств Homoptera для склонового экотона хребта Абишира-Ахуба.
Для склонового экотона особым является преобладание в отряде Orthoptera (рис. 28) особей, находящихся на личиночной стадии развития, что объясняется более поздним онтогенезом этих особей. Тем не менее, выделено семейство Tettigoniidae – c видом Decticus verriucivorus всего лишь 13 % от общего количества особей.

Рис. 28. Структура населения семейств Orthoptera для склонового экотона хребта Абишира-Ахуба.
При рассмотрении антропогенной нагрузки в пределах экотонного участка хребта Абишира-Ахуба, следует опереться на соотношение антропофитной и разнотравно-злаковой растительных ассоциаций (рис. 29).

Рис 29. Соотношение антропофитной и разнотравно-злаковой растительности в пределах склонового экотона хребта Абишира-Ахуба.
Из диаграммы (рис. 29) следует, что антропофитами занято 65% района исследования, а значит антропогенная нарушенность составляет 40-50% территории. Выше рассмотренные участки не идут ни в какое сравнение с интенсивной антропогенной нагрузкой на Абишира-Ахубе, где ведется не только чрезмерный выпас скота, но и мало ограниченная вырубка лесов. Здесь, безусловно, повсеместно встречается антропофитная растительность (Конева, 2005). Также индикаторами нарушенности являются и насекомые семейства Chrysomelidae, которые «привязаны» к антропофитам – щавелю конскому, крапиве двудомной, чемерице Лоббиля и др.
Однако невозможно не отметить активное и продолжительное антропогенное воздействие на биоценозы экотонов, при котором соотношения тепла и влаги превышают экологически допустимые для видов пределы, нарушает функциональные связи, обеспечивающие буферный механизм организации ландшафтной структуры. Это приводит к значительной дестабилизации географической среды экотонов, о чем свидетельствуют многочисленные экологически кризисные и катастрофические ситуации на экотонных территориях. Такие ситуации не только определили повышенное внимание к экотонам экологов и географов, но и их успешное сотрудничество, в том числе на международном уровне (Экотоны …, 1997).
В настоящее время антропогенное воздействие человека на природные ландшафты возрастает, что приводит к деградации почв, растительного покрова, смене естественных ассоциаций антропофитной растительностью, изменению состава хортобионтов, особенно в зоне высокогорного экотона субальпийских лугов. Поэтому остро встает вопрос о необходимости изучения этих процессов.
Широкое распространение экотонных территорий подтверждает их значительную роль в поддержании гомеостаза биосферы, а процесс антропогенной экотонизации охватывает территорию, значительно превышающую площадь очагов воздействия.
Выводы по главе.
При общем сравнении отрядов хортобионтов по всем видам экотона можно сказать, что на всех участках доминирует отряд Homoptera, только для хребта Абишира-Ахуба преобладает отряд Hemiptera, который является следующим по преобладанию среди беспозвоночных. Но насекомые этих отрядов находились на личиночной стадии развития в долине Птыша, причем более 70% особей. Для хребтовых районов исследования особи были во взрослом состоянии, но ввиду отсутствия специалиста - энтомолога, к сожалению более детально не удалось определить видовую или родовую принадлежность. Так как развитие у клопов проходит с неполным превращением, то личинки их обычно похожи на взрослых и по внешнему виду, и по образу жизни, что также осложняет определение.
Но насекомые многочисленны и способны адаптироваться к сложной для обитания среды. Большинство клопов развивается в году в одном поколении. Зимовка происходит обычно во взрослой фазе или (например, у клопов-слепняков) в фазе яйца.
Среди видов семейства прямокрылых одинаковых видов не выявлено, к тому же они не отличаются ни численными поазателями, ни видовым многообразием.
Самыми богатыми по видовому сочетанию явился отряд Coleoptera, хотя он не отличается большим количеством. Но доминирует этот отряд среди всех видов экотона на водораздельном, а минимально – на ледниковом. Достаточное количество особей прямокрылых представлено личинками. Полностью взрослые особи отмечены только на Малой Хатипаре. А на Абишира-Ахубе и в долине Птыша более 80% насекомых представлены личинками. В целом, различия на видовом уровне беспозвоночных характерны как для видов экотона, так и для фаций водораздельного экотона.
Более детально проанализируем видовую структуру семейств отряда Coleoptera. Характерными для всех трех видов экотона явились семейство Curculionidae – вид Zacladus geranii, причем общими явились и рода Otiorhynchus и Nastus, а также семейство Chrysomelidae – род Cryptocephalus.
Общими для ледникового и водораздельного экотонов явились семейство Cantharidae с видом Rhagonicha limbata при доминировании разных видов – Cantharis sudetica и Rhagonicha translucida – соответственно. При сравнении семейства Coccinellidae – общим видом явился Coccinula quatuordecimpunctata.
Характерными и для склонового, и для водораздельного видов экотона явились семейство Elateridae – род Athous sp.(доминирует на Малой Хатипаре) и вид Dascillus cervinus (ярко выражен на Абишира-Ахубе) и семейство Oedemeridae – вид Oedemera flavescens.
Значит возможно проведение закономерности по выявлению связи между геоморфологическим, климатогенным факторами и формированием растительности и распространением беспозвоночных хортобионтов.
Заключение
В последние десятилетия в экологии стало выделяться особое направление, связанное с исследованием сообществ беспозвоночных животных, обитающих в травянистом ярусе растительности. Травостой рассматривается как самостоятельный ярус животного населения суши; комплекс его обитателей - специальный объект синэкологических исследований с применением особых методик. В настоящий момент предлагается назвать обособляющееся направление хортозоологией. Животную компоненту сообществ называют "животным населением". В широком смысле население - совокупность особей различных видов, обитающих на определенном пространстве.
Толща травостоя обладает своеобразным микроклиматом. Для беспозвоночных в силу их мелких размеров и особенностей физиологии огромное значение имеет именно микроклимат, определяющий условия их жизни. В однородных, казалось бы, условиях небольшого участка луга на самом деле микроклиматические характеристики сильно различаются в зависимости от того, в какой точке произвести замеры.
В толще травостоя при продвижении от вершин растений к поверхности почвы наблюдается падение освещенности, что связано со сложными процессами отражения, поглощения и прохождения солнечных лучей зелеными листьями. Важное значение имеет густота травостоя, общее проективное покрытие почвы, пространственное расположение листовых пластинок.
В ходе проведенного исследования в пределах зоны экотона субальпийского пояса была изучена и апробирована методика сбора полевого материала при помощи энтомологического сачка, собрана коллекция насекомых по трем основным группам – Orthoptera, Coleoptera, Hemiptera и Homoptera.
В итоге на внутриландшафтную дифференциацию насекомых оказывают влияние многие факторы. В значительной степени сказывается высота, значительное увлажнение территории и уровень антропогенной нагрузки. Водораздельный экотон является эталонным в плане сохранности природного компонента. Склоновый экотон сильно подвержен антропогенному фактору. Ледниковый экотон только проходит стадии заселения территории растительными ассоциациями, беспозвоночными.
В результате исследования данного участка была проведена сравнительная характеристика основных групп насекомых травяного покрова. Соответственно, наиболее сильные различия наблюдались в зоне экотона, что связано с различиями переходной зоны
В заключении следует отметить, что данная работа позволяет четко представить распределение насекомых в пределах трех видов экотона субальпийского пояса высокогорий, а также дифференциацию внутри него на уровне фаций водораздельного хребта.
Литература
| 1. | Асланикашвили Н.А., Мамулян М.А. Влияние ориентации горных хребтов Кавказа на направление и скорость ветра // Тр. Зак. НИГМИ. 1990. Вып 89 (96). С. 188-198. |
| 2. | Арманд Д. Л. Наука о ландшафте. – М., 1975. |
| 3. | Атаев З.В. Ландшафтная структура Предгорного Дагестана / Антропогенные ландшафты: структура, методы и прикладные аспекты изучения. - Воронеж: ВГУ, 1988. |
| 4. | Бей-Биенко Г.Я. Прямокрылые – Orthoptera и кожистокрылые –Dermaptera // Животный мир СССР. Т. 5. – М.; Л.: АН СССР, 1958.– С. 435-456. |
| 5. | Белоновская Е.А. Структура экотона верхней границы леса на Северном Кавказе // Биота экосистем Большого Кавказа. М., Наука, 1990. – С. 6-40. |
| 6. | Берчашвили Н.Л. Кавказ: ландшафты, модели, эксперименты. -Тбилиси, 1995. |
| 7. | Беручишвили Н.Л. Сезонная динамика структуры и функционирования фаций // Ландшафтный сборник.- Тбилиси. - 1979. - С. 100-115. |
| 8. | Братков В.В. Ландшафтно-геофизический анализ природно-территориальных комплексов Северо-Восточного Кавказа. Автореферат канд. диссертации. - Тбилиси, 1992. |
| 9. | Братков В.В., Салпагаров А.Д., Мокроусов Д.О. Сезонная динамика ландшафтов Тебердинского заповедника / Труды Тебердинского заповедника. Вып. 41. М. – Ставрополь, 2005. |
| 10. | Братков В.В., Салпагаров Д.С. Ландшафты Северо-Западного и Северо-Восточного Кавказа. – Москва: Илекса; Ставрополь: Ставропольсервисшкола, 2001. – 256с. |
| 11. | Волынкин И.Н., Доценко В.В. Ландшафты и физико-географическое районирование Чечено-Ингушетии / Проблемы физической географии Северо-Восточного Кавказа. - Грозный, 1979. |
| 12. | Волынкин И.Н., Доценко В.В. Структура ландшафтов Чечено-Ингушетии / Природа и хозяйство восточной части Северного Кавказа.- Орджоникидзе, 1977. |
| 13. | Второв П.П. Биоэнергетика и биогеография некоторых ландшафтов Терскей Ала-Тоо. – Фрунзе: Илим, 1968. – 167 с. |
| 14. | Гвоздецкий Н.А. Кавказ. Очерк природы. - М., 1963. – 264с. |
| 15. | Гельмгольц Н.Ф. Горно-долинная циркуляция северных склонов Тянь-Шаня. Л.: Гидрометеоиздат. 1963. 330 с. |
| 16. | Герасимов И.П. Экологические проблемы в прошлой, настоящей и будущей географии мира. – М.: Наука, 1985. |
| 17. | Герасимов И.П., Лилиенберг Д.А. Геоморфологическая модель Большого Кавказа // Большой Кавказ – Стара Планина (Балканы). М.: Наука, 1984.- с.9-38. |
| 18. | Геренчук К.И., Миллер Г.П., Трохимчук С.В. О морфологической структуре горных ландшафтов / Материалы к VI Всесоюзному совещанию по вопросам ландшафтоведения. - Алма-Ата, 1963. |
| 19. | Горностаев Г.Н. Насекомые СССР. М.: Мысль, 1970. – 372 с. |
| 20. | Грегори К. География и географы. Физическая география. – М.: Прогресс, 1988. – 384 с. |
| 21. | Дьяконов К.Н. Ландшафтная политика в современном обществе // География на рубеже тысячелетий. Труды XII съезда Русского географического общества. Т. 1. – СПб., 2005. - С. 105-106. |
| 22. | Ефремов Ю. В., Ильичев Ю. Г., Панов В. Д., Панова С. В., Погорелов А.В., Шереметьев В. М. Хребты Большого Кавказа и их влияние на климат. - Краснодар: Просвещение-Юг, 2001. - 145 с. |
| 23. | Ефремов Ю. В., Панов В. Д., Ильичев Ю. Г. Рельеф и современные экзогенные процессы в верховьях рек Теберды, Малый и Большой Зеленчук // Оценка экологического состояния горных и предгорных экосистем Кавказа. Вып. 3. Ставрополь,2000. – с. 84-94. |
| 24. | Ефремов Ю.В., Ильичев Ю.Г., Панов В.Д., Панова С.В., Погорелов А.В., Шереметьев В. М. Хребты Большого Кавказа и их влияние на климат. – Краснодар.: Просвещение-Юг, 2001. - 145 с. |
| 25. | Исаченко А.Г. Ландшафт географический // БСЭ. – 3-е изд. – М., 1973. - Т.14. –144с. |
| 26. | Исаченко А.Г. Ландшафтоведение и физико-географическое районирование. – М.: Высшая школа, 1991. – 365 с. |
| 27. | Исаченко А.Г. Ландшафтоведение и физико-географическое районирование. – М.: Высшая школа, 1990. |
| 28. |
Исаченко А.Г. Основные проблемы ландшафтоведения горных стран // Вопросы ландшафтоведения. – Алма-Ата, 1972. – С. 6-22. Копанева Л.М. Прямокрылые (Orthoptera) Главного Кавказского хребта в верховьях реки Теберды и их вертикальное распределение // Зоол. журн, 1962. Т. XVI. |
| 29. | Исаченко А.Г. Основы ландшафтоведения и физико-географическое районирование. – М.: Высшая школа, 1965. |
| 30. | Кавказ. Природные условия и естественные ресурсы СССР. М.: Наука. 1966. 482 с. |
| 31. | Коломыц Э.Г. Ландшафтная организация зонального географического пространства и его границ // Изв. РАН сер. геогр., 1996 – №2. – С. 39-57. |
| 32. | Конева В.В. Влияние антропогенных факторов на внутриландшафтную дифференциацию насекомых в зоне экотона (на примере хребтов М. Хатипара и Абишира-Ахуба). // Международной научно-практической конференции “Проблемы экологической безопасности и сохранения природно-ресурсного потенциала” (29-30.09.2006) Ставрополь, 2005. |
| 33. | Конева В.В. Внутриландшафтная дифференциация насекомых хребта Малая Хатипара. // Вопросы физической географии. Материалы 50-й научно-методической конференции «Университетская наука - региону» СГУ. – Ставрополь: Изд-во СГУ, 2005. |
| 34. | Конева В.В. Особенности внутриландшафтной дифференциации хортобионтов хребта М. Хатипара в зоне экотона субальпийского пояса (на уровне фаций). // Вопросы физической географии. Материалы 52-й научно-методической конференции «Университетская наука - региону» СГУ. – Ставрополь: Изд-во СГУ, 2007. |
| 35. | Кочуров Б. И. Геоэкология: экодиагностика и эколого-хозяйственный баланс территории. – Смоленск: СГУ, 1999. – 154 с. |
| 36. | Крыжановский О.Л., Тер-Минасян М.Е. Жесткокрылые – Coleoptera // Животный мир СССР. Т. V. – М.; Л.: АН СССР, 1958. – С 384-429. |
| 37. | Лавренко Е.М. Типы вертикальной поясности растительности в горах СССР / Современные проблемы географии, 1964. |
| 38. | Лиховид А.А., Шальнев В.А., Шкарлет К.Ю., Харин К.В., Юрин Д.В., Разумов С.И., Маслиев, А.В. О биотике ландшафта, итогах и перспективах развития // Труды XII съезда Русского Географического Общества. Том 2, СПб, 2005. |
| 39. | Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. Определитель насекомых Европейской части СССР. - М.: Просвещение, 1970. 304с. |
| 40. | Мамаев Б.М. и др. Определитель насекомых европейской части СССР. - М.: Просвешение, 1976. – 304 с. |
| 41. | Мильков Ф.Н. Ландшафтная география и вопросы практики. – М.: Мысль, 1966. – 364 с. |
| 42. | Мильков Ф.Н. Физическая география: учение о ландшафте и географическая зональность. – Воронеж, 1986. |
| 43. | Михайлов Н.И. Физико-географическое районирование. Курс лекций, ч. 2. – М.: МГУ, 1962. |
| 44. | Михеев Г.А., Снежко Г.А., Сафронов И.Н. Геологическое строение территории Архызского участка Тебердинского заповедника // Тр. Тебердинского заповедника. 1962. Вып. IV. С. 37-55. |
| 45. | Нееф Э. Теоретические основы ландшафтоведения. – М., 1974. – 218 с. |
| 46. | Николаев В. А. Ландшафтные экотоны //Вестн. Моск. ун-та. Сер. 5. Геогр. 2003., 6. |
| 47. |
Олейникова Д.В. Внутриландшафтная дифференциация травянистой растительности высокогорных ландшафтов Западного Кавказа: Автореф. Дисс канд. Географ. наук. – Ставрополь: Изд-во СГУ, 2005. |
| 48. | Онищенко В.В., Салпагаров А.Д. Геоэкологические признаки и предпосылки сезонного развития растительности Северо-Западного Кавказа (на примере Тебердинского заповедника) / Тр. ТГБЗ. Вып. №32. – М.: НИА-Природа, 2002. – 186с. |
| 49. | Определитель насекомых европейской части СССР в пяти томах. – М.; Л.: Изд-во Наука, 1964. |
| 50. | Преображенский В.С. Острые проблемы ландшафтоведения на рубеже веков // Вестник МГУ, серия географическая, 1998, № 3. |
| 51. | Программа и методика биогеоценологических исследований / Под ред. Дылис Н.В. М.: 1974 – С. 57-59. |
| 52. | Савельева В.В. Ландшафты Архыза // Северный Кавказ. Вып. 2.(Вопросы физической и исторической географии), Ставрополь 1973. – С. 42-54. |
| 53. | Сафронов И.Н. Геоморфология Северного Кавказа. Изд-во Ростовского ун-та, 1960. – 216 с. |
| 54. | Сочава В. Б. Введение в учение о геосистемах. – Новосибирск: Наука, 1978. |
| 55. | Сочава В.Б. География и экология. // Материалы V съезда ГО СССР. – Л.: 1970. |
| 56. | Тайлаков Т.А. Об условиях образования фена над Северным Кавказом // Тр. ГМЦ. 1967. Вып. 6. С. 120-127. |
| 57. | Темникова Н.С. Климат Северного Кавказа и прилегающих степей. Л.: Гидрометеоиздат, 1959. – 367 с. |
| 58. |
Тумаджанов И.И. Кавказские луга и криволесья // Растительность европейской части СССР. – Л., 1963. – С. 198-202. |
| 59. | Тушинский Г. К. Геоморфологический очерк Тебердинского государственного заповедника. Тр. ТГЗ, т 1., 1957. |
| 60. | Тушинский Г.К. Современное и древнее оледенение Тебердинского района // Ежегодник Советского альпинизма – М., 1949. |
| 61. | Хрусталев Ю. П. Эколого – географический словарь. – Батайск, Изд-во Рост. ун-та,2000. |
| 62. |
Чернов Ю.И., Руденская Л.В. Об использовании энтомологического кошения как метода количественного учёта беспозвоночных -обитателей травяного покрова // Зоол. жур., т. XLIX, вып. 1, 1970. – С. 12-14. |
| 63. | Черных В.И. Гравитационная тектоника и ее отражение в рельефе западной части Лабино-Малкинской зоны // Тезисы докладов 7 краевой конференции по геологии и полезным ископаемым Северного Кавказа. Ессентуки, 1991. – С. 39-48. |
| 64. | Чупахин В.М. Физическая география Северного Кавказа. – Ростов н/Д, РГУ, 1974. |
| 65. | Шальнев В. А. Ландшафты хребта Малая Хатипара / Сев. Кавказ – вып. 2. Вопросы физической и исторической географии. Ставрополь.: СГПИ, 1973. |
| 66. | Шальнев В. А. Особенности теплового баланса луговых ассоциаций // Изв. ВГО. – Т. 103. – Вып. 2., 1971 (б). |
| 67. | Шальнев В. А. Опыт выделения морфологических единиц горных ландшафтов в пределах северного склона Главного Кавказского хребта. Спб. ”Вопросы географии Сев. Кавказа”. Ставрополь, 1971. |
| 68. | Шальнев В. А. Оценка роли экспозиции склонов в формировании фаций горных стран // Изв. ВГО. Т. 103. Вып. 3., 1971 (а). |
| 69. | Шальнев В. А., Джанибекова Х. А. Ландшафты Карачаево - Черкессии // Вестник СГУ. – Вып. 6, 1996. |
| 70. | Шальнев В. А., Чикалин А. Н. Некоторые особенности радиационного режима луговых ассоциаций // Изв. ВГО, т 100, вып. 2, 1968. |
| 71. | Шальнев В. А., Чикалин А. Н. Особенности градиентных различий основных метеорологических элементов луговых ассоциаций // Изв. ВГО. – Т. 100. – Вып. 4., 1971. |
| 72. | Шальнев В. А., Чикалин А. Н. Особенности градиентных различий основных метеорологических элементов луговых ассоциаций // Изв. ВГО. – Т. 100. – Вып. 4., 1968 (а). |
| 73. | Шальнев В. А., Юрин Д. В. Ландшафтный подход в изучении экотона горных территорий // Вестник СГУ. – Вып. 12. 1997. |
| 74. | Шальнев В.А. Ландшафты Домбая // Вопросы физической географии. Материалы 50-й научно-методической конференции «Университетская наука - региону» СГУ. – Ставрополь: Изд-во СГУ, 2005. |
| 75. | Шальнев В.А. Ландшафты Северного Кавказа: эволюция и современность. – Ставрополь: Изд-во СГУ, 2004. – 165с. |
| 76. | Шальнев В.А. Современные проблемы моделирования ландшафта //Вестник Ставропольского гос. университета. 1999. № 17. – С. 23-28. |
| 77. | Шальнев В.А., Кондратьева А.А. Экосистемный подход в изучении горных ландшафтов (на примере долины р. Учкулан) // Рукоп. деп. ВИНИТИ № 2026-В-94.08.1994. |
| 78. | Шальнев В.А., Конева В.В. Внутриландшафтная дифференциация насекомых высокогорного долинно-ледникового экотона (на примере реки Птыш). // Вопросы физической географии. Материалы 51-й научно-методической конференции «Университетская наука - региону» СГУ. – Ставрополь: Изд-во СГУ, 2006. |
| 79. | Шальнев В.А., Чикалин А.Н. Радиационный режим луговых ассоциаций хребта М. Хатипара / Тр. Тебердинского гос. заповедника. Т. VIII. – Ставрополь, 1971. |
| 80. | Шальнев В.В., Нефедова М.В. // Экотон в морфологии горных ландшафтов (на примере Бокового хребта Северо-Западного Кавказа). Материалы XI ландшафтной конференции, - М., 2006. |
| 81. | Шкарлет К. Ю., Разумов С. И. Особенности внутриландшафтной дифференциации животного населения высокогорий (на примере хребта Малая Хатипара) // Вопросы физической географии. – Ставрополь, СГУ, 2003. – 123 с. |
| 82. | Щукина О. Е. О климатических факторах формирования ландшафтной поясности в горных странах // Изв. ВГО. – Т. 92. – Вып. 1, 1960. |
| 83. | Виноградов Б. В. Основы ландшафтной экологии. - М., ГЕОС. - 1994. - 418 с. |
| 84. | Милановский Е.Е., Хаин В.Е. Геологическое строение Кавказа. М., 1963. – 355 с. |
| 85. | Фасулати К.К. Полевое изучение наземных беспозвоночных. Изд. 2. М.: Высшая школа, 1971. – 424 с. |
| 86. | Экотоны в биосфере. / Под ред. В. С. Залетаева. - М., 1997. - 329 с. |
| 87. | Clements F.E. Plant succession and indicators. - New York, 1928. 453p. |
| 88. | Troll C. Luftbildplan und oekologische Bodenorschung // Zeitschr. Geos/ Erdkunde za Berlin. 1939, № 7, №8. |
| 89. | Vink A.P., 1983. Landscape ecology and land use. London, Londman. |
| 90. | http:// www.bionet.ru |
| 91. | http://flora-world.by.ru/ Publikations/Kniga_3/Tez_1. htm. |
| 92. | http://www.floranimal.ru/orders/2712/ html. |
| 93. | http://zoo ex.baikal.ru. |
| Билеты и ответы по туризму и экскурсиям | |
|
БИЛЕТ № 1 Школы научного управления: общая характеристика, значение для сферы сервиса и туризма. Порядок образования туристской фирмы. Туристский ... По горным склонам террасами спускаются в долину реки Жэхэ крестьянские поля, издалека виднеются черепичные крыши храмов, многоярусные пагоды и величественные силуэты горных вершин. В горах наблюдается вертикальная поясность со сменой растительного покрова: остепненные участки в долинах сменяются по склонам широколиственными и хвойными лесами, альпийскими ... |
Раздел: Рефераты по туризму Тип: реферат |
| Русский остров | |
|
Содержание Введение Глава 1. Географическое положение Глава 2. Климатические особенности Глава 3. Ландшафт Глава 4. Растительность Глава 5. Животный ... В днищах речных долин и ложков структура растительности отвечает режиму обводнения. Влажные типы дубовых лесов занимают пологие склоны, высокие речные террасы и вершины долин горных ключей. |
Раздел: Рефераты по физкультуре и спорту Тип: курсовая работа |
| Кавказская горная страна | |
|
... КАЗАХСКИЙ НАЦИОНАЛЬНЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ им. АБАЯ КУРСОВАЯ РАБОТА На тему "Кавказская горная страна" Выполнила: Дыгаева Замира ГОЭР-3 Он отличается от предыдущего появлением внизу сухих степей и у самого подножия полупустынь; резким сужением лесной зоны (на внешних склонах хребтов горного Дагестана) и развитием у ... Основной части Джавахетско-Армянского нагорья свойствен переднеазиатский тип структуры высотной зональности ландшафтов с зонами полупустынь; горных степей, нагорных ксерофитов и ... |
Раздел: Рефераты по географии Тип: курсовая работа |
| Заповедники Казахстана | |
|
Содержание. Введение 2 Физико-географическая характеристика заповедников Казахстана 3 Природные парки 8 Природные заказники 9 Физико-географическое ... Основные типы охраняемых ландшафтов - горные склоны Курчумского хребта, озеро Маркаколь с наклонной приозерной равниной, пересеченной несколькими горными речками, самой крупной из ... Заказник занимает восточную часть хребта Заилийского Алатау на востоке 2000-4000 м. Здесь преобладают ландшафты среднегорий и высокогорий с субальпийскими лугами, на значительной ... |
Раздел: Рефераты по географии Тип: реферат |
| Ландшафтно-экологические методы исследований | |
|
СОДЕРЖАНИЕ ВВЕДЕНИЕ 1. ОСНОВНЫЕ ЗАДАЧИ И МЕТОДЫ ФИЗИЧЕСКОЙ ГЕОГРАФИИ 1.1 Основные задачи физической географии. Этапы научного познания Основные задачи ... Примерами подурочищ могут служить покатый прибалочный склон северной экспозиции, сложенный с поверхности покровными суглинками, с серыми лесными средне- и тяжелосуглинистыми ... Макроформы имеют площадь от сотен до десятков тысяч квадратных километров (например, хребты и впадины горной страны, возвышенности и низменности на равнине, долины крупных рек). |
Раздел: Рефераты по географии Тип: учебное пособие |