Реферат: Регуляция клеточного деления и скорости роста клеток
Регуляция клеточного деления и скорости роста клеток
Существует понятие о клеточном цикле – последовательности событий от одного деления клетки до другого. Клеточный цикл прокариотической и эукариотической клеток различается весьма существенно. Учитывая большую сложность организации клеток эукариот, проще начать с рассмотрения механизмов регуляции процессов клеточного деления и роста клеток прокариот, тем более что в биотехнологических процессах все большее распространение получает культивирование эукариотических клеток с использование подходов, применяемых для культивирования одноклеточных прокариот.
Последовательность событий в процессе деления клетки
Процесс клеточного деления у прокариот включает следующие события в определенной очередности:
1) накопление «критической» клеточной массы;
2) репликация ДНК генома;
3) построение новой клеточной оболочки;
4) построение клеточной перегородки;
5) расхождение дочерних клеток.
Некоторые из этих событий протекают одновременно, другие строго последовательно или вообще могут отсутствовать.
Регуляция клеточного деления складывается из регуляции каждого из этих событий и организации их взаимодействия, при котором в клеточном делении устанавливается последовательность процессов и вырабатываются сигналы для инициации следующего по порядку процесса.
Накопление критической клеточной массы и репликация ДНК
Это необходимые подготовительные этапы собственно клеточного деления. Следует отметить, что размер клеток каждого микроорганизма, растущего сбалансированно в стандартных условиях, является достаточно постоянной величиной, чтобы служить одним из таксономических признаков. В.Д. Донаши даже ввел понятие элементарной клетки, т.е. наименьшей, возможной для данного микроорганизма. Таким образом, существуют механизмы, включающие процесс деления клетки при накоплении ее пороговой массы.
Построение новой клеточной оболочки
Необходимо различать пролиферацию цитоплазматической мембраны и клеточной стенки и сегрегацию поверхностных структур.
При изучении пролиферации используют, как правило, синхронные культуры микроорганизмов и изучают включение меченных радиоизотопами соединений путем равновесного или импульсного введения этих соединений.
Таким путем установлено, что включение белков в цитоплазматическую мембрану Escherichia coli и Bacillus subtilis следует сложной кинетике, свидетельствующей о запасании предобразованных белков в цитоплазме, в период подготовки клеточного деления и быстрой их мобилизации – в процессе построения клеточной перегородки. В период деления возрастает активность некоторых литических ферментов, участвующих в образовании «брешей» в предсуществующем каркасе клеточной стенки, необходимых для включения новых ее фрагментов. Таким образом, регуляция активности этих ферментов осуществляется путем временного перевода их в скрытое состояние с последующей мобилизацией в необходимый момент. Точных данных о механизмах такой регуляции нет, но можно полагать, здесь имеет место взаимодействие ферментов с мембранами.
При изучении сегрегации поверхностных слоев также используют введение в эти структуры меченых предшественников с прослеживанием их судьбы через несколько генераций после переноса клеток на среду, не содержащую метки. Наблюдения обычно осуществляют методом электронно-микроскопической радиоавтографии, где в качестве метки используется тритий, который в силу небольшой энергии р-частиц дает на радиоавтографах короткие треки, удобные для определения мест локализации метки.
Другой подход – наблюдение за образованием и распределением маркеров структурных компонентов оболочки в течение нескольких генераций после их индукции. В этом случае удобно использовать специфические маркеры клеточной стенки или цитоплазматической мембраны, или, наконец, такие общие маркеры, как жгутики.
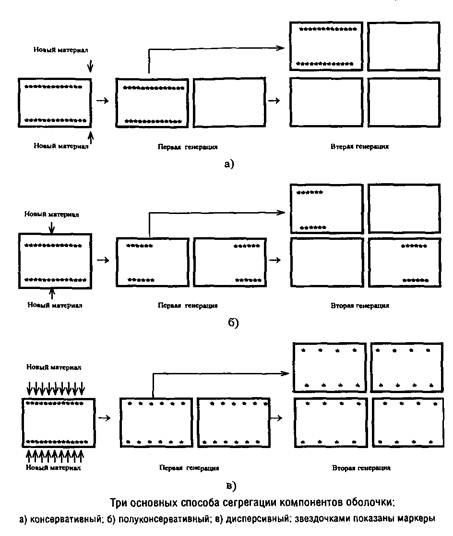
Можно представить себе три основных способа локализации сайтов включения предшественников: консервативный, полуконсервативный и дисперсивный. В первом случае после второй генерации лишь четверть клеток содержит маркеры, во втором случае – половина клеток, а в третьем – все клетки.
Вопрос о механизме сегрегации поверхностных слоев можно считать более или менее однозначно решенным лишь для кокковидных форм бактерий в случае, если они характеризуются мономорфным клеточным циклом и делятся в одной плоскости. Для этих форм разные экспериментальные подходы дают сходную картину, указывающую на полуконсервативный способ сегрегации. Для палочковидных форм бактерий сведения о способе сегрегации противоречивы.
Однозначное установление локализации мест включения мембранных компонентов затрудняется их значительной латеральной подвижностью, составляющей, например, для липополисахарида наружной мембраны Escherichia coti около 1 мкм за 25 с. Кроме того, способ сегрегации может определяться скоростью роста микроорганизма: у медленно растущих клеток Escherichia coii он близок к биполярному, а у быстро растущих становится днсперсивным.
Построение клеточной перегородки
В изучении механизмов регуляции данного этапа клеточного цикла важную роль сыграли специфические мутанты, особенно мутанты Escherichia colt и Bacillus subtilis, образующие миниклетки-мутанты). Миниклетки возникают на полюсах нормальных клеток, имеют небольшие размеры и не содержат хромосомной ДНК. Однако у них нормальный аппарат транскрипции и трансляции, поэтому они могут быть использованы для изучения функционирования захваченных из материнской клетки плазмид, а также введенных извне искусственных генетических элементов, полученных методами генетической инженерии. Именно существование т/л-мутантов позволило сделать вывод, что сайт, ответственный за образование перегородки и локализующийся в процессе деления в экваториальной зоне клетки, сохраняется на полюсах дочерних клеток. В норме эти полярные сайты выключаются и могут функционировать наряду с вновь формирующимися экваториальными сайтами лишь у mm‑мутантов.
В любой из клеток т/л-мутанта одновременно существуют два функционально активных сайта построения перегородки, но в клеточном цикле срабатывает лишь один из них.
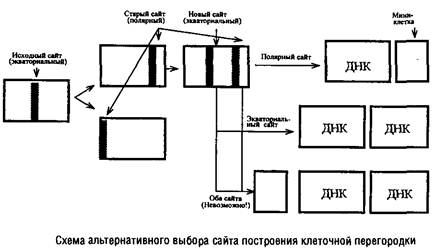
Оказалось невозможным образование одновременно трех клеток: двух нормальных и одной мини. Поэтому сделан вывод о существовании определенного компонента – активатора сборки клеточной перегородки. По-видимому, на протяжении клеточного цикла образуется ограниченное количество этого активатора, достаточное для функционирования лишь одного сайта, и оно полностью расходуется в этом процессе.
Обнаружить существование такого кванта в нормальных клетках невозможно, так как количество квантов активатора и количество функционирующих сайтов у них совпадает, а у т/л-мутантов это количество превышает число квантов активатора.
Характер взаимосвязи процессов клеточного деления
Между процессом накопления критической массы клетки, репликацией ДНК и построением клеточной перегородки не обнаружено облигатно-реципрокной связи, при которой подавление одного из процессов тормозило бы другие и наоборот. Например, в случае Bacillus subtitis возможно построение перегородки и формирование клеток нормального размера после подавления репликации ДНК налидиксовой кислотой. В результате одна из дочерних клеток не содержит ДНК. Кстати, такие клетки, не содержащие ДНК, нечувствительны к пенициллину, вызывающему лизис только активно растущих клеток, поэтому данный антибиотик можно использовать для получения их чистой популяции без ДНК для дальнейших исследований.
Можно получить и обратную картину, если ингибировать построение клеточной перегородки низкими концентрациями пенициллина G. Аналогично действует повышение температуры в случае некоторых л-мутантов. При этом рост клетки и репликация ДНК могут продолжаться, приводя к возникновению «многонуклеоидных» нитей, которые после удаления ингибитора фрагментируются на соответствующее число нормальных клеток.
Замечено, что клеточный цикл прокариот, таких как Escherichia coli, при росте на минеральной среде с глюкозой можно подразделить на два основных периода. Они получили обозначения периодов Си D. Иногда в периоде D выделяют еще период Т – время от появления первых признаков клеточной перегородки до завершения клеточного деления.
Период С в норме занимает около 40 мин., фактически представляя собой время полной репликации генома Escherichia coli, которое мало зависит от скорости роста. В последнем случае инициация нового цикла репликации ДНК происходит до завершения клеточного деления, и дочерние клетки получают уже частично реплицированную ДНК, так что к моменту деления репликация успевает завершиться.
Период D занимает около 20 мин. – между моментом завершения репликации и моментом окончательного формирования клеточной перегородки.
Для нормального протекания клеточного цикла необходимо, чтобы в период С происходила не только репликация ДНК, но и синтез белка и РНК, так как ингибиторы транскрипции и трансляции, введенные в течение периода С, тормозят клеточное деление и увеличивают время генерации. Если же ввести эти ингибиторы на период, не превышающий 15 мин., деление клетки завершается вовремя. Очевидно, что минимальная длительность периода D может быть равна периоду Т, т.е. времени, необходимому для сборки перегородки. Эти выводы подтверждаются фактом, что данные ингибиторы, введенные в период D, не тормозят клеточное деление. Следовательно, предшественники, необходимые для построения клеточной перегородки, и другие белки, важные для завершения деления клетки, синтезируются в период С и хранятся в резерве до начала сборки перегородки.
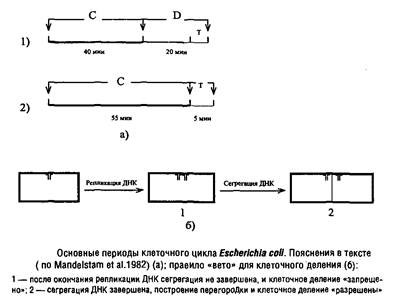
Центральное место в проблеме регуляции клеточного деления занимает вопрос о природе сигнала, необходимого для запуска процесса сборки клеточной перегородки. Длительное время считалось, что этим сигналом является терминация репликации ДНК, однако рассмотренные нами свидетельства, указывающие на отсутствие облигатной связи между этими процессами, делают подобное заключение сомнительным.
Недавно установлено, что подавление сегрегации вновь синтезированных цепей ДНК, достигаемого в период D за счет сборки клеточной оболочки из предшественников, препятствует завершению клеточного цикла. Поэтому можно полагать, что для нормального построения клеточной перегородки от ДНК должен быть освобожден сайт, ответственный за сборку перегородки, локализованный в экваториальной части клетки и занятый ДНК сразу после завершения ее репликации. Отсюда вывод: регуляторное взаимодействие между репликацией ДНК и построением клеточной перегородки состоит в своеобразном правиле «вето» со стороны ДНК. Если нарушен процесс нормальной сегрегации реплицированной ДНК и соответствующее место в экваториальной области клетки занято, то сборка клеточной перегородки не может быть осуществлена и клеточное деление тормозится. Формально в этом случае наблюдается зависимость между репликацией ДНК и делением клетки.
Взаимодействие регуляторных механизмов при управлении скоростью роста микроорганизмов
Один из узловых вопросов, связанных с управлением скоростью роста микроорганизмов – о механизмах перестройки метаболизма микробной клетки при изменении состава питательной среды.
В хемостатной культуре регулирование состава среды позволяет получить клетки определенного химического состава, а иногда и с заранее заданными свойствами. Например, для получения клеток, обогащенных белком, но со сниженным содержанием нуклеиновых кислот целесообразно использовать лимитирование по фосфору.
При обогащении среды, допустим, путем добавления дополнительных питательных веществ, а в хемостатной культуре путем увеличения протока среды, скорость роста увеличивается до нового значения, которое, как правило, не является максимально возможным в силу неполной реализации потенциала клетки. Это происходит из-за наличия так называемых узких мест, т.е. биохимических реакций, ограничивающих скорость всего процесса, а выявляя их, можно получить максимальный выход биомассы и ценных для человека продуктов метаболизма.
Таблица 1. Влияние различных видов лимитирования на состав клеток микроорганизма (типа Escherichia coli)
| Лимитируемый источник | Состав клеток | |||
| Белок | Нуклеиновые кислоты | Липиды | Запасные вещества | |
| Углерод | Не влияет | Не влияет | Снижает | Снижает |
| Азот | Снижает | Снижает | Повышает | Повышает |
| Фосфор | Не влияет или повышает | Снижает | Снижает | Повышает |
| Цинк | Не влияет | Снижает | Не влияет | Не влияет |
Рассмотрим значение разных уровней регуляции, представленных на схеме, для управления общей скоростью роста организма.
Обычно скорость транспорта субстратов более или менее точно сбалансирована со скоростью их метаболизма, а иногда превышает ее. В последнем случае в клетке формируется резерв субстратов, способный оказывать разнообразное, в том числе тормозящее, действие на метаболизм клетки, если отсутствует трансрегуляторное ингибирование транспорта этих субстратов из среды их внутриклеточным пулом. При некоторых условиях транспорт оказывается лимитирующим этапом метаболизма, например при дефиците в среде необходимых субстратов и кофакторов, особенно в случае организмов, не способных к синтезу данных веществ или осуществляющих эти процессы с пониженной скоростью. Аналогичная ситуация создается при недостаточной эффективности транспортных систем, даже если в среде избыток субстрата. Этап выделения продуктов может лимитировать рост, если продукт обладает ингибиторным или отрицательным регуляторным действием на метаболизм. В клетке при этом может вырабатываться специальный механизм для активного удаления таких веществ.
В тех случаях, когда транспортный процесс становится узким местом, лимитирующим общую скорость метаболизма, воздействие, активирующее транспорт или повышающее избирательную проницаемость клеточной оболочки, может положительно влиять на скорость роста организма. Этап функционирования ферментов может оказаться рост-лимитирующим звеном метаболизма лишь при отсутствии в клетке необходимого количества фермента. При этом быстро включаются компенсирующие механизмы: наступает индукция фермента или снимается репрессия его синтеза. Для конститутивных ферментов возможна стимуляция на уровне трансляции. Только при недостаточной эффективности всех этих регуляторных механизмов количество фермента может оказаться неадекватным условиям роста.
Во многих случаях несбалансированного роста наиболее вероятными претендентами на роль «узких мест» метаболизма являются процессы синтеза макромолекул, особенно РНК и белка. Этап репликации редко выступает в качестве узкого места метаболизма, хотя скорость элонгации ДНК – величина достаточно постоянная, составляющая у Escherichia coli около 2000 пар нуклеотидов в секунду, и мало зависит от условий выращивания. Это объясняется специальной организацией регуляторных механизмов, настроенных таким образом, что при улучшении условий питания повышается частота инициации новых циклов репликации ДНК. Поэтому, если время генерации меньше, чем период репликации ДНК, то новые циклы репликации инициируются до завершения старых и в быстро растущих клетках ДНК присутствует в виде сильно разветвленной структуры, соответствующей по массе 3–8 эквивалентам генофора. При этом, очевидно, локусов, расположенных вблизи от точки начала репликации, в клетке значительно больше, чем расположенных ближе к точке терминации, что может вызывать повышение синтеза некоторых белков. Однако чаще всего эффект дозы гена не проявляется из-за регуляции на уровне транскрипции и трансляции.
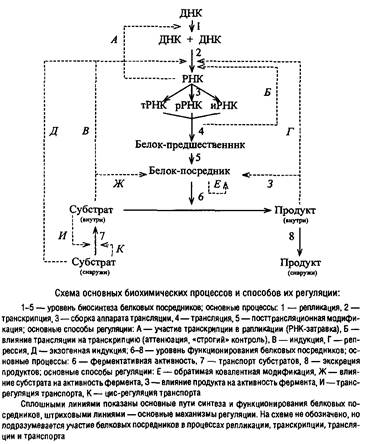
Менее определенной оказывается ситуация с транскрипцией. Длительное время считалось, что скорость элонгации в транскрипции является такой же постоянной величиной, как и в репликации. Но появляется все больше сведений о том, что в транскрипции она может варьировать.
Существует тесное сопряжение между элонгацией РНК в процессе транскрипции и элонгацией полипептидной молекулы в процессе трансляции, и выражается оно не только в пространственном сопряжении процессов, как это имеет место при аттенюации, но и в регуляторном воздействии через молекулы эффекторов. Торможение элонгации трансляции приводит к синтезу специфического эффектора гуанозинтетрафосфата, который существенно влияет на процесс транскрипции.
Дефицит энергии также тормозит гидролиз ppGpp, так как активность пирофосфатгидролазы АТР-зависима. Таким образом, при аминокислотном голодании не только стимулируется синтез PpGpp, но и тормозится его гидролиз.
Кроме этого механизма, по-видимому, существует еще один путь синтеза ppGpp, так как при дефиците источников энергии он накапливается даже в клетках мутанта Escherichia coli. У некоторых бацилл и стрептомицетов установлен фактор, независимый от рибосом, катализирующий синтез ppGpp при снижении уровня АТР в клетке. Накопление ppGpp в клетках приводит к резкому торможению образования стабильных форм РНК и, соответственно, к торможению формирования аппарата трансляции, избыточное количество которого в условиях голодания становится излишним и даже вредным. Это и есть так называемый строгий контроль. Одновременно подавляется транскрипция локусов рибо-сомных белков и факторов элонгации трансляции. Однако ppGpp оказывает и положительное действие на транскрипцию: он стимулирует транскрипцию некоторых аминокислотных регулонов, а также регулонов азотного метаболизма.
Кроме влияния на транскрипцию ppGpp регулирует активность ряда ключевых ферментов метаболизма, участвующих в образовании нуклеотидов, фосфолипидов, пептидогликана, в транспорте азотистых оснований и т.д. Наконец, ppGpp активирует некоторые протеолитические системы клетки, ускоряя внутриклеточный протеолиз.
Все изложенное делает понятной необходимость тонкой регуляции уровня ppGpp в клетке.
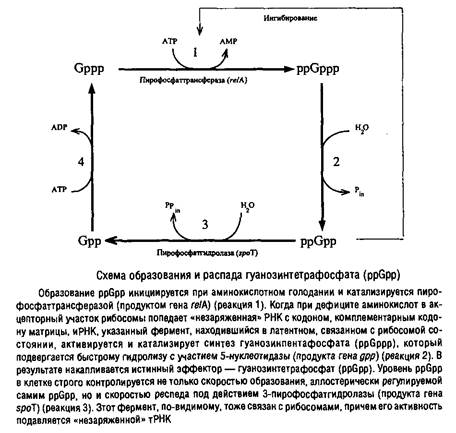
Необходимо отметить, что гуанозинполифосфаты аналогичного или иного строения обнаружены в клетках многих про- и эукариот, где они выполняют различные регуляторные функции.
Таким образом, сопряженный процесс транскрипции-трансляции оказывается во многих случаях решающим этапом приспособления клетки к условиям голодания, например при переносе на бедную среду.
При обратной ситуации – переносе клеток на богатую среду (shift-up) именно процессы сопряженной транскрипции-трансляции являются наиболее узким местом метаболизма, лимитирующим общую скорость роста популяции.
После обогащения среды происходит «вспышка» синтеза белка, тРНК переходит в «заряженное» состояние, в результате резко снижается образование ppGpp и запускается быстрый синтез стабильных форм РНК, чему способствует множественная репрессия ранее функционировавших оперонов, В итоге синтез белка и скорость роста увеличиваются, пока позволяет сопряженное функционирование процессов транскрипции-трансляции.
Из всего изложенного следует практический вывод, касающийся селекции и конструирования штаммов-продуцентов, способных к «сверхсинтезу» ценных продуктов. Например, для стимуляции синтеза аминокислот образование ppGpp оказывается полезным, поэтому более перспективными продуцентами могут оказаться штаммы Ret. Напротив, конструирование штаммов, образующих белковые продукты, предполагает необходимость подавления внутриклеточного протеолиза, что требует использования штаммов Ret или других условий, подавляющих образование ppGpp.
| Ответы на вопросы по генетике | |
|
1. Предмет генетики и ее связь с другими науками Генетика, как самостоятельная наука выделилась из биологии в 1900 году. Термин генетика введён в 1906 ... Синтез ДНК - репликация ДНК - процесс самоудвоения ДНК. В период репликации ДНК и образования перегородки клетки непрерывно растёт, идёт формирование рибосом и других соединений. |
Раздел: Рефераты по биологии Тип: шпаргалка |
| Цели регенеративной медицины. Терапия стволовыми клетками | |
|
Содержание 1. Терапия стволовыми клетками 2. Типы стволовых клеток 2.1 Эмбриональные стволовые клетки 2.2 Гемопоэтические стволовые клетки 2.3 ... Высказано предположение, что независимо от их линейного происхождения, стволовые клетки отвечают сходным образом на регуляторные сигналы самообновления и дифференцировки, и похоже ... Относительный состав оснований ДНК регуляторных последовательностей недетерминированных полипотентных родоначальных клеток может иметь значение для частоты транскрипции этих генов ... |
Раздел: Рефераты по медицине Тип: курсовая работа |
| Мононуклеарный онкогенез | |
|
МОНОНУКЛЕАРНЫЙ ОНКОГЕНЕЗ Введение злокачественный мононуклеарный заболевание лечение Онкогенез (греч. onkos - масса, нарост, опухоль; genesis ... Структурные изменения клеточной мембраны и химические изменения в цитоплазме клетки, влияют на проявление генотипических изменений ядерной ДНК - эпигенетический механизм. Вирхов Р. (1863-1867): хроническое раздражение тканей вызывает с их стороны защитную реакцию - идет регенерация и скорость деления клеток увеличивается, но если будет утерян ... |
Раздел: Рефераты по медицине Тип: книга |
| Физиология растений | |
|
Куниченко Наталья Александровна, кандидат сельскохозяйственных наук, доцент, заведующая кафедрой защиты растений и экологии Приднестровского ... Калий участвует в регуляции тургора клетки, избирательной проницаемости клеточных мембран. Эта группа гормонов не обладает столь ярко выраженным действием, как предыдущие, однако цитокинины воздействуют на многие звенья метаболизма, усиливают синтез ДНК, РНК, белков. |
Раздел: Рефераты по биологии Тип: учебное пособие |
| Клетка | |
|
Реферат по Биологии Тема: "Клетка" Исполнил: Лежнин Пётр 818 гр. -2001- ВВЕДЕНИЕ Цитология - наука о клетках - элементарных единицах строения ... Коренное улучшение всей техники микроскопирования позволило исследователям к началу XX столетия обнаружить основные клеточные органоиды, выяснить строение ядра и закономерности ... Митохондрия - это органоид клетки, в котором вырабатывается основная масса энергии клетки, сконцентрированная в АТФ и используемая затем в разнообразных процессах синтеза и во всех ... |
Раздел: Рефераты по биологии Тип: реферат |