Контрольная работа: Строение и состав живой клетки
Введение
Строение и состав живой клетки
Введение
Все живые организмы состоят из клеток и подразделяются на одноклеточные и многоклеточные. Популяция одноклеточных организмов включает клетки одного типа или небольшого количества типов, тогда как у многоклеточных организмов клетки специализированы. По типу строения клетки подразделяются на прокариотческие и эукариотические.
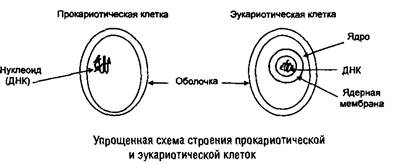
Все клеточные организмы имеют сходный химический состав и содержат три основных типа макромолекул: ДНК, PHK белки, а также полисахаридные и липидные компоненты. Кроме того, в клетках присутствуют переменные количества низкомолекулярных веществ — субстратов и продуктов энергетических и конструктивных процессов (аминокислоты, сахара, нуклеотиды и др.). В среднем около 80% клеточной массы составляет вода.
1. Клеточные стенки и клеточные мембраны
Как правило, клетки окружены двумя оболочками: клеточной стенкой и цитоплазматической мембраной, для эукариотических клеток применяют название плазмолемма.
Клеточная стенка обеспечивает механическую прочность клетки, придавая ей жесткую (ригидную) структуру, благодаря чему клетка выдерживает высокое внутреннее осмотическое давление (5—20 МПа). Кроме того, клеточная стенка может обусловливать некоторую степень избирательной проницаемости для низкомолекулярных веществ, а также способность взаимодействовать с другими клетками, вирусами и физическими поверхностями. Строение клеточной стенки у разных организмов имеет свои особенности.
Клетки большинства тканей многоклеточных животных не содержат выраженной клеточной стенки. Растительные клетки, напротив, имеют очень сложную клеточную стенку, построенную из целлюлозных микрофибрилл, погруженных в матрикс (из пектина и гемицеллюлоз).
Клеточные стенки дрожжей и мицелиальных грибов состоят из гомо- и гетерополисахаридов (глюканов, хитина) и белкового комплекса, выполняющего антигенную роль. Толщина этих слоев достигает 1 мкм.
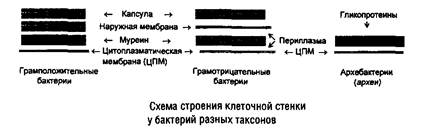
Особенности клеточных стенок бактерий связаны со способностью бактерий окрашиваться по Граму. Клеточная стенка громположительных бактерий построена в основном из гетерополисахарида муреина (пептидогликана), содержащего аминокислотные «мостики». В состав клеточной стенки входят также тейхоевые кислоты (рибит- или глицеринтейхоевые), ковалентно связанные с мурамовой кислотой пептидогликана. У грамотрицательных бактерий слой муреина невелик, но в клеточной стенке присутствует наружная мембрана, построенная из фосфолипидов, белков и липопо-лисахарида, обеспечивающая некоторую степень избирательной проницаемости и содержащая рецепторы фагов и антигены. Толщина клеточной стенки составляет от 15 до 80 нм. Существуют бактерии, полностью лишенные клеточной стенки (микоплазмы). У представителей архебактерий (архей) в клеточной стенке отсутствует муреин (иногда содержится отличающийся по составу псевдомуреин), а осмопротекторную роль выполняет слой гликопротеинов (рис. 3).
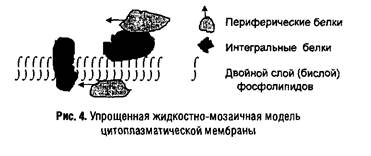
Цитоплазматическая мембрана состоит из белков и липидов в соотношении от 1: 4 (в миелине, мембране нервных волокон) до 4:1 (в мембранах бактерий). Во многих мембранах присутствуют небольшие количества углеводов (до 5%) и следы РНК, а также неорганические катионы (в основном Са2+ и Mg2+). Существенную часть мембран составляет вода — она участвует в формировании гидрофобных (энтропийных) связей между компонентами мембраны. Еще в 1930 г. Д. Даниэлян предложил модель мембраны в виде двойного липидного слоя или бислоя. Развитием этих представлений является жидкостно-мозаичная модель, предложенная С.Д. Сингером и Г. Николсоном в 1972 г., согласно которой мембранные белки подразделяются на периферические (слабо связанные с мембраной) и интегральные (локализованные в липидном бислое). Последние свободно плавают в «липидном море», перемещаясь в латеральном направлении (в плоскости мембраны), но неспособны «пронырнуть» на противоположную ее сторону. Таким образом, мембраны оказываются асимметричными, по крайней мере, в отношении белкового состава. Асимметрия наблюдается и в расположении липидов в мембране, но менее выражена. Некоторые из интегральных белков имеют трансмембранную ориентацию и выполняют функции переносчиков субстратов или сигналов. Схема подобной мембраны представлена на рис. 4.
2. Состав мембранных липидов
Основная часть мембранных липидов представлена фосфолипидами, в основе которых лежит глицерин-3-фосфат. Такие фосфолипиды называют фосфоацилгпицеринами. В их молекуле гидроксильные группы глицерина при Cf и С2 этерифицированы жирными кислотами, а остаток фосфорной кислоты либо остается свободным, как в диацилглицерин-3-фосфате (или фосфатидной кислоте), либо этерифицирован спиртовыми гидроксилами серина, этаноламина, холина, инозита, глицерина или глицерина и лизина. Соответственно они называются: фосфатидилсерин, фосфатидилэтаноламин (кефалин), фосфатидилхолин (лецитин), фосфатидилинозит, фосфатидилглицерин, лизилфосфатидилглицерин. Объединение молекул фосфатидной кислоты и фосфатидилглицерина дает дифосфатидилглицерин (кардиолипин), обладающий иммунологическими свойствами (рис. 5).

Удаление одной из ацильных групп приводит к лизофосфолипидам, которые обладают выраженными поверхностно-активными свойствами и могут, например, способствовать эмульгированию жиров в кишечнике.
Мембраны грамположительных бактерий содержат все эти фосфолипиды (кроме лецитина), тогда как у грамотрицательных бактерий основным, а то и единственным мембранным фосфолипидом является фосфатидилэтаноламин.
Соотношение различных классов фосфолипидов в мембране существенно зависит от условий выращивания организма. Например, при снижении рН среды в мембранах бактерий начинают преобладать положительно заряженные фосфолипиды (аминоацилфосфатидилглицерины, фосфатидилэтаноламин), наличие которых существенно уменьшает проницаемость мембраны для протонов. В большинстве случаев фосфолипиды представляют собой сложные эфиры глицерина или других многоатомных спиртов, но существуют липиды, в которых одна из спиртовых групп глицерина образует простую эфирную связь с ал килами или алкенами. Необычные липиды найдены у большой группы прокариот, так называемых архебактерий (или архей), в качестве простых эфиров глицерина и длинноцепочечных С-20, С-40-гидроизопреноидных спиртов (полипренолов), точнее, дифитанилдиглицериновых диэфиров или дибифитанилдиглицериновых тетраэфиров (рис. 6).
Мембрана, образованная тетраэфирами, уже не может рассматриваться как двойной липидный слой в истинном значении этого понятия и не подвергается расщеплению по гидрофобной сердцевине методом «замораживания-скалывания» (freeze-fracture). Она более устойчива к стрессовым воздействиям окружающей среды, что позволяет микроорганизмам, имеющим такие мембраны, существовать в экстремальных условиях рН и температуры (экстремофилы). У термофилов дополнительная стабилизация достигается формированием в полипренольной цепи пятичленньгх циклов, уменьшающих вращательную подвижность углеводородных цепей.

Кроме фосфолипидов, построенных на основе глицерина, в клетках эу- и прокариот встречаются фосфолипиды, которые являются производными диодов: этиленгпиколя, 1,2- и 1,3-пропандиолов, 1,3-, 1,4- и 2,3-бутандиолов, а также 1,5-пентандиола. Среди диольных липидов встречаются моно- и диацильные производные — это сложные эфиры различных жирных кислот; простые эфиры, смешанные алкильные (или алкенильные) и ацильные производные, диольные аналоги фосфатидилхолина, фосфатидилэтаноламина и др. Обычно содержание диольных липидов составляет только 0,5—1,5% содержания глицериновых липидов. Но некоторые морские моллюски и иглокожие содержат в тканях примерно равные количества производных глицерина и этиленгликоля. Содержание диольных липидов уменьшается в течение зимы, поэтому они предположительно могут использоваться как запасные вещества. Замечено также их увеличение в процессе регенерации печени у крыс.
Другим классом «безглицериновых» липидов являются сфинголипиды, основу которых составляет алифатический аминоспирт сфингозин (или дигидросфингозин) (рис. 7). Построенные на его основе церамиды широко распространены в тканях растений и животных, но количество их незначительно. Они обнаружены также в пилях Escherichia coli. Сфингомиелины входят в состав нервной ткани, липидов крови и некоторых других компонентов клеток животных.

В большинстве биологических мембран содержатся также гликолипиды. В клетках животных они, как правило, являются производными сфингозина, у которого к первичному гидроксилу присоединен остаток сахара или олигосахарида. Если сахарами являются глюкоза или галактоза, такие липиды называются цереброзидами (особенно много их в тканях мозга). Если углеводной частью является олигосахарид, речь идет о ганглиозидах (выделены из ганглий, мозга и ряда других тканей). Ганглиозиды найдены также в пилях некоторых бактерий (Neisseria) и определяют прилипание этих патогенов к поверхности клеток животного организма. У прокариот (в основном у грамположительных бактерий и цианобактерий) гликолипиды также содержат глюкозу, галактозу и маннозу. Их количество в мембранах невелико, но при лимитировании фосфором может возрастать, при этом гликолипиды, по-видимому, замещают фосфолипиды. Наконец, в мембранах большинства эукариот (а также у Mycoplasma) содержатся стероиды, в основном холестерин (у животных), эргостерин (у дрожжей) и стигмастерин (у растений). Микоплазмы неспособны синтезировать стерины, но требуют присутствия их в среде для стабилизации клеточной мембраны.
3. Природа и состав жирных кислот в мембранных липидах
Природа жирных кислот в липидах мембран зависит как от вида организма, так и от условий его существования. Наиболее часто встречающиеся жирные кислоты липидов животных, растительных и прокариотных клеток.
С увеличением числа двойных связей значительно снижается температура плавления жирных кислот (а также содержащих эти кислоты липидов) и повышается их растворимость в неполярных растворителях. Поскольку функциональная активность мембранных белков регулируется фазовым состоянием липидов мембраны (как правило, в жидком состоянии их активность выше), при снижении температуры в мембране должно повышаться содержание ненасыщенных кислот. Благодаря постоянству внутренних условий (гомеостазу) животного макроорганизма влияние температуры на жирнокислотный состав липидов обычно проявляется слабо, но, например, в липидах нижних конечностей пингвинов повышено содержание ненасыщенных жирных кислот.
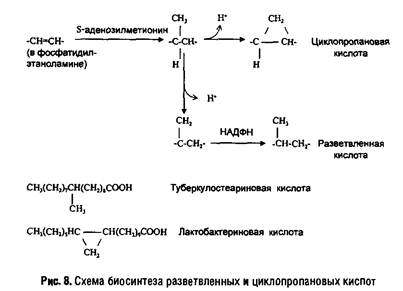
В мембранах прокариот разнообразие жирных кислот довольно велико. Особенностью бактерий является наличие разветвленных и циклопропановых кислот. У некоторых грамположительных бактерий (например, Micrococcus luteus) 90% жирных кислот липидов составляют разветвленные кислоты, а ненасыщенные кислоты практически отсутствуют. Грамотрицательные бактерии содержат смесь насыщенных и ненасыщенных кислот с преобладанием С-16 и С-18 кислот, а также циклопропановые кислоты (например, лактобацилловую кислоту). Пути их биосинтеза показаны на рис. 8.
Изменение температуры существенно влияет на соотношение насыщенных и ненасыщенных жирных кислот в мембранах прокариот, при этом снижение температуры культивирования приводит к увеличению доли ненасыщенных кислот. Предельными случаями являются психрофилы, растущие при температурах, близких к 0"С, у которых присутствуют практически только ненасыщенные жирные кислоты, а также термофилы (привычная температурная среда выше 60°С) — у них все жирные кислоты липидов насыщенные.
Тем не менее состав жирных кислот фосфолипидов у бактерий при культивировании в стандартных условиях достаточно постоянен, и его можно использовать как таксономический признак — в интересах классификации или при анализе состава микробных сообществ без изолирования в чистых культурах отдельных представителей.
Для изучения структуры, состава и функции органелл необходимо, как правило, изолировать их из клетки в чистом виде. Обычно это достигается методом дифференциального центрифугирования после разрушения клеток в механических или ультразвуковых гомогенизаторах.
Кратко охарактеризуем основные процессы и органеллы (в алфавитном порядке).
Аэросомы — однослойные везикулы, мембрана которых построена только из белка. Способствуют повышению плавучести клеток, так как в них содержится газовая фаза, совпадающая по составу с газовой фазой окружающей среды.
Вакуоли — мембранные образования, служащие для поддержания тургорного давления, запасания различных веществ, а также выполняющие лизосомные функции.
Внехромосомная ДНК. В митохондриях и хлоропластах содержится ДНК, образующая нуклеоид бактериального типа. Заключенная в ней генетическая информация не дублируется в ядерной ДНК и способна к автономному выражению в белках посредством собственных систем транскрипции и трансляции (включающих рибосомы 70 S бактериального типа).
У прокариот внехромосомная ДНК организована в виде плазмид, которые могут существовать и реплицироваться автономно или в интегрированном в хромосому состоянии (например, в виде профага).
Гидрогеносомы — окружены однослойной мембраной и содержат комплекс пируватдегидрогеназ (у трихомонад).
Гистоны — представляют собой положительно заряженные (основные) белки, входящие в состав хромосом в комплексе с ДНК (обнаружены также у архебактерий).
Гликосомы — окружены однослойной мембраной и содержат ферменты гликолиза (у некоторых протозойных микроорганизмов, в частности у возбудителей сонной болезни).
Глиоксисомы — разновидность пероксисом — место локализации ферментов глиоксалатного шунта, участвующих в превращении запасных жиров в углеводы. Поэтому они тесно ассоциированы со сферосомами, жирозапасающими органеллами растительных клеток.
Жгутики и реснички представляют собой аппарат, определяющий подвижность клеток или их способность создавать поток окружающей среды к органам поглощения пищи. Жгутики эукариот и прокариот сильно различаются по строению и составу.
Карбоксисомы — окружены однослойной мембраной и содержат ключевые ферменты фиксации углекислоты в цикле Кальвина (у фототрофных и некоторых хемолитотрофных прокариот).
Лизосомы — мембранные везикулы, содержащие гидролитические ферменты, участвующие в круговороте белков, полисахаридов, липидов и нуклеиновых кислот. Для предотвращения неорганизованного действия этих ферментов они заключены в органеллу, окруженную однослойной мембраной. Некоторые наследственные болезни связаны с недостаточностью лизосомных ферментов.
Микротрубочки и микрофиламенты, по-видимому, исполняют роль цитоскелета и формируются из белка тубулина. Они входят в состав центриолей, играющих важную роль в делении ядра, а также в состав жгутиков и ресничек.
Митоз — деление ядра, сопровождающееся удвоением числа хромосом; мейоз — деление ядра без удвоения числа хромосом, в результате чего образуются гаплоидные клетки.
Митохондрии являются местом осуществления окислительных процессов, в них локализованы ферменты цикла трикарбоновых кислот, дыхательная цепь, система окислительного фосфорили-рования. В клетках содержится от 1 до нескольких тысяч митохондрий. Митохондрии содержат внехромосомную ДНК, способны к самовоспроизведению и, возможно, ведут свое начало от прока-риотических клеток (эндосимбиотическая гипотеза). Однако часть белков этих органелл кодируется ядерными генами, поэтому они не культивируются автономно. У прокариот сходные по функциям структуры, происходящие из цитоплазматической мембраны, называют мезо сомами.
Пероксисомы — органеллы, окруженные однослойной мембраной и содержащие ряд окислительных ферментов, а также каталазу, разрушающую перекись водорода, образующуюся в процессе окисления. Могут рассматриваться как более древние аналоги митохондрий и играют важную роль в обмене липидов.
Пили, фимбрии представляют собой выросты поверхности клеток и бывают простые и половые. Простые пили построены из белка пилина и играют важную роль в прикреплении бактерий к субстрату. На поверхности клетки их может быть 50—400 штук. Половые пили (1 на клетку) используются в процессе конъюгации. Выросты клеток, содержащие цитоплазму, называют простеками.
Они служат для увеличения поверхности клеток, прикрепления к субстрату или участвуют в почковании клеток.
Хитосомы — место локализации хитинсинтетазы, встречаются у грибов. Осуществляют функцию переноса микрофибрилл хитина к клеточной стенке.
Хлоропласты содержат хлорофилл и осуществляют процесс фотосинтеза у растений. Как и в митохондриях, в них содержится собственная ДНК. Аналогичные по функциям бактериальные структуры называют хроматофорами, а также тилакоидами и хлоросомами. В отличие от хлоропластов они не содержат ДНК и происходят из цитоплазматической мембраны.
Целлюлосомы — содержат комплекс ферментов и липидов. Определяют присоединение бактерий к целлюлозным субстратам и их расщепление.
Эндоплазматтеский ритикулум — это внутриклеточная сеть взаимосвязанных, ограниченных мембраной трубочек и пузырьков. Мембрана эндоплазматического ретикулума образует единое целое с ядерной мембраной. Если на наружной поверхности эндоплазматического ретикулума адсорбированы рибосомы, его называют шероховатым, если нет — гладким. Шероховатый эндоплазматический ритикулум — место синтеза секретируемых белков, которые попадают внутрь его каналов, а затем поступают в «цистерны» аппарата Гольджи (диктиосомы у растений), откуда переносятся внутри мембранных везикул либо в плазмалемму, либо в мембрану вакуолей (тонопласт) и секретируются внутрь вакуолей или в окружающую среду (экзоцитоз).
Ядро (нуклеоплазма) эукариотических клеток окружено двойной мембраной (плазмолеммой), содержащей поры (поросомы). В ядре присутствует ядрышко (место синтеза РНК и сборки рибосом). В отличие от ядра нуклеоид прокариотических клеток не отделен мембраной от цитоплазмы и представляет собой комплекс ДНК, РНК и белков.
| Педиатрия | |
|
1. История отечественной педиатрии Начало становления отечественной педиатрии относится к 18 веку. Ломоносов в письме к Шувалову указывает на ... Значительно активизируется перекисное окисление липидов, в крови накапливаются свободнорадикальные соединения, повреждающие клеточные мембраны и нарушающие процессы обмена на ... В результате нарушается проницаемость клеточных мембран, уменьшается синтез кальцийсвязывающего белка, обеспечивающего транспорт ионов кальция через кишечную стенку, снижается ... |
Раздел: Рефераты по медицине Тип: шпаргалка |
| Клетка | |
|
Реферат по Биологии Тема: "Клетка" Исполнил: Лежнин Пётр 818 гр. -2001- ВВЕДЕНИЕ Цитология - наука о клетках - элементарных единицах строения ... Хотя вирусы, как и клетки, состоят из двух основных компонентов - нуклеиновой кислоты и белка, структура вирусов и клеток резко различна, что не позволяет считать вирусы клеточной ... В состав мембран аппарата Гольджи входят липиды, или, точнее, фосфолипиды и белки. |
Раздел: Рефераты по биологии Тип: реферат |
| Все о клетке | |
|
Реферат по Общей биологии На тему: "Клетка" Ученицы 11 "В" класса Физико-математической школы № Преподаватель: Москва 2001 год План реферата: Введение ... Клетки содержат множество разнообразных органических соединений: углеводы, липиды, белки, нуклеиновые кислоты и др. Фосфолипиды в состав молекул, которых входит остаток фосфорной кислоты, являются основой всех клеточных мембран. |
Раздел: Рефераты по биологии Тип: реферат |
| Перспективы применения липосомальных форм | |
|
Оглавление Введение Глава 1. История открытия липосом Глава 2. Применение липосом в медицине 2.1 Свойства липосомальных частиц 2.2 Использование ... Формы взаимодействия липосом с мембраной клетки: липосома может увеличить проницаемость мембраны - вызвать образование дополнительных каналов (I); может прикрепиться к мембране ... Небольшой объем водного раствора, содержащего вещество А, отделен от окружающей среды бислойной липидной мембраной, состоящей из соевого или яичного лецитина с включением других ... |
Раздел: Рефераты по медицине Тип: дипломная работа |
| Акушерство и гинекология | |
|
акушерство и гинекология (лекции) ОГЛАВЛЕНИЕ I АКУШЕРСТВО 1. Общие понятия в акушерстве 2. Физиология беременности 3. Гестозы 4. Родовой акт 5 ... Развивается спазм и других сосудов ° гипоксия, в условиях которой активируется ПОЛ°продукты пероксидации повреждают клеточные мембраны°повышается проницаемость клеточных мембран ... Клотримазол - вызывает торможение синтеза нуклеиновых кислот, липидов, полисахаридов клетки гриба, что в свою очередь приводит к повреждению клеточной оболочки и увеличению ... |
Раздел: Рефераты по медицине Тип: учебное пособие |