Курсовая работа: Белки клеточного цикла в отделах мозга сусликов citellus undulatus на разных стадиях гибернационного цикла
Федеральное агентство по образованию
Пензенский государственный педагогический университет
им. В. Г. Белинского
Факультет Естественно-географический
Кафедра биохимии
БЕЛКИ КЛЕТОЧНОГО ЦИКЛА В ОТДЕЛАХ МОЗГА СУСЛИКОВ CITELLUS UNDULATUS НА РАЗНЫХ СТАДИЯХ ГИБЕРНАЦИОННОГО ЦИКЛА
Введение
Зимняя спячка представляет собой форму адаптации животных к действию неблагоприятных условий обитания: низкой температуры окружающей среды, отсутствию пищи и т.д. [6, 39, 4] Затрачивают все уровни организации от биохимического до поведенческого, и реализуются генетические программы, запускаемые при действии соответствующих внешних (укорочение фотопериода, понижение температуры, бескормица и т.д.) и внутренних факторов (цирканнуальные ритмы нейромедиаторов, пептидов и гормонов) [ 3, 32].
Состояние гибернации у зимоспящих животных характеризуется тем, что при переходе в это состояние температура тела понижается и часто достигает околонулевых значений. Наблюдается существенное снижение частоты сердечных сокращений – приблизительно с 250 до 8-10 ударов в 1 мин и снижение частоты дыхания – со 150 до одного или менее дыхательных циклов в 1 мин [ 5 ]. Отмечается снижение уровня метаболизма от 0,2 кал/(г·мин-1) при гибернации [ 1 ]. Показано, что общее потребление энергии в сезон гибернации (включая периодические регулярные выходы из спячки) у гибернирующих животных на 87% меньше, чем у активных [ 39 ]. В течение гибернационного сезона наблюдаются периодические выходы животных из состояния спячки с соответствующим повышением уровня метаболизма и температуры тела [ 39 ]. Таким образом, гибернация делится на периоды спячки (бауты), которые прерываются короткими (обычно 24 ч или меньше) периодами эутермии.
Процессы погружения в спячку и пробуждения, а также длительность баутов контролируются центральной нервной системой. Даже во время глубокой зимней спячки некоторые отделы мозга сохраняют достаточно высокую активность. Так, в лимбической системе (а именно в септуме, гиппокампе, гипоталамусе) наблюдается почти постоянная ЭЭГ-активность на протяжении всего баута [ 10 ].
Проблема зимней спячки имеет не только общебиологический интерес, но и привлекает внимание с непосредственно практической точки зрения: полученные фундаментальные данные могут быть положены в основу разработки методов создания гипобиоза, управляемого при помощи естественных механизмов, что чрезвычайно важно для клиники, когда требуется поддержать организм более или менее длительное время в состоянии значительно сниженных физиологических функций. Такая регулируемая искусственная гипотермия, вызывающая лишь умеренно вредные эффекты, способствует повышению устойчивости организма к влиянию ряда неблагориятных факторов, прежде всего тканевой гипоксии, которая часто сопутствует различным видам патологических процессов, в частности нарушениям кровообращения [ 3 ]. Результаты исследования нейрогенеза у сусликов при смене функционального состояния мозга во время гибернационного цикла в литературе отсутствуют, однако наличие его показано у птиц при смене сезонов и наступлении весны. Ряд авторов рассматривает гибернацию как модель ишемии мозга, при которой снижение кровотока, доставки кислорода и глюкозы во время спячки, а также последующая реоксигенация при выходе из спячки не приводят к повреждению и гибели клеток. Однако для ряда ишемических состояний, связанных с нейродегенерацией и гибелью клеток мозга, показано увеличение интенсивности нейрогенеза в постишемический период.
В связи с этим, целью работы явилось исследование экспрессии белков клеточного цикла (циклинов – А, В1, циклинзависимых киназ- cdc2, cdk2, cdk 4, МСМ2) и не связанной с клеточным циклом циклинзависимой киназы cdk5 в мозге сусликов Citellus undulatus на разных стадиях гибернационного цикла.
Задачи:
1. Определить экспрессию белков клеточного цикла:
· циклинов- ClnA, ClnB1;
· циклинзависимых киназ- cdc2, cdk2, cdk4;
· маркёра пролиферирующих клеток MCM2;
2. Определить экспрессию циклинзависимой киназы cdk5, не связанной с клеточным циклом.
3. Выявить различия в экспрессии белков клеточного цикла на разных стадиях гибернационного цикла и при разной температуре тела животных.
Глава 1. Обзор литературы
1.1 Биохимические изменения в тканях при зимней спячке
Зимняя спячка (гибернация) – это закрепленная в ходе эволюции уникальная способность к минимализации жизненных функций организма, позволяющая ряду видов млекопитающих в течение многих месяцев переживать холод, бескормицу, сокращения светлого периода суток [ 5 ]. В Европе, Азии, Америке впадают в зимнюю спячку почти все виды летучих мышей, ежи и многие грызуны: сони, тушканчики, хомяки, бурундуки, суслики и сурки [ 6, 39 ].
Основой этого приспособления, по-видимому, являются крайне разнообразные пути сохранения энергетического баланса организма при сезонных изменениях во внешней среде. Подсчитано, что в течение зимней спячки (7-8 мес.) сусликов экономится от 80-90% энергии, которая могла бы быть потрачена на поддержание обычного уровня метаболизма [ 34 ].
Зимняя спячка является прерывистым процессом и состоит из баутов. Баут спячки состоит из входа в гипобиоз, пребывания в торпидном состоянии и выхода из него [39]. Каждый баут спячки может продолжаться от нескольких часов до нескольких недель и зависит как от физиологических особенностей животных, так и условий окружающей среды [ 37 ].
Температура тела млекопитающих во время зимней спячки снижается вплоть до 0°С. Понижение температуры тела приводит не только к частичной или полной потере чувствительности и двигательной активности, но и подавлению активности всех функций и систем организма: кровообращения, дыхания, пищеварения, водно-солевого обмена, гормонального статуса, размножения [37, 39]. Характерным для спячки является резкое замедление дыхания, в то время как в активном состоянии животное делает от нескольких десятков до нескольких сот дыханий в минуту [6]. Выход из состояния оцепенения сопровождается резким усилением дыхания, быстрой реперфузией тканей, активацией липолиза [36]. Известно, что уровень потребления кислорода отражает уровень метаболизма. Так во время пробуждения за 2 часа температура тела повышается с 0°С до 37°С, а потребление кислорода увеличивается в 50 раз [ 27, 36 ].
У гибернирующих животных сильно (более чем на 50%) увеличивается масса тела [34]. Известно, что при гибернации жир млекопитающих становится основным энергетическим субстратом [12]. Обильные запасы жировой ткани обеспечивают животным существование в течение длительного времени [34].
Бауты гибернации длиннее и глубже в середине зимы. В периоды пробуждения животные разогреваются и просыпаются. Температура тела приходит в норму [34, 35].
Пробуждение млекопитающих происходит или вследствие согревания при повышении температуры среды, или при понижении ее ниже 0°С.
Скорость пробуждения различна, но обычно оно происходит очень быстро. Резко увеличивается частота дыхания и сердцебиения. Повышается интенсивность окислительных процессов в организме, почти полностью расходуется запас гликогена печени [6]. При пробуждении, в фазе повышенного термогенеза, частота сердцебиений достигает более 400 уд/мин [9 ]. Ведущая роль в регуляции зимней спячки и периодических пробуждений принадлежит нервной и эндокринной системам [1]. У гибернирующих животных наряду с биохимическими наблюдаются и некоторые морфологические изменения в ткани мозга. Так, древовидные структуры дендритов укорачиваются при понижении температуры тела и быстро растут при её повышении; при впадении в спячку сокращается площадь тела нейрона [18, 19 ].
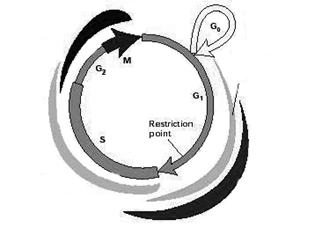
1.2 Клеточный цикл
В норме клеточный цикл - это последовательность событий, приводящих к делению исходной клетки на две дочерние. Его подразделяют на четыре фазы (или стадии): S фаза (в ней происходит репликация ДНК) и М фаза (фаза митоза) разделены промежутками – это фазы G1 и G2. Покоящиеся клетки находятся в G0 фазе [30 ].
Для нервных клеток млекопитающих показано, что повторное вступление клетки в клеточный цикл зависит от внеклеточных сигналов. К ним относят митогенные стимулы (например, факторы роста FGF и EGF, тромбин, сыворотка) и факторы дифференцировки (цитокины, стрессорные влияния на клетку – такие как ультрафиолетовое излучение, или повреждающие ДНК воздействия) [ 31 ] . Эти сигналы осуществляют свое влияние через последовательную экспрессию, активацию и деактивацию так называемых белков клеточного цикла - комплексов циклинов (Cn) с циклин-зависимыми протеинкиназами (cdk). Cn/cdk комплексы обеспечивают развитие клеточного цикла, и тонко регулируются множеством механизмов: транскрипцией, фосфорилированием, протеолизом, а также активностью ингибиторов циклин-зависимых протеинкиназ (i-cdk). В ходе онтогенеза экспрессия и активное взаимодействие циклинов, Cn/cdk комплексов и i-cdk сопровождается выходом клетки из клеточного цикла, приводящим к дифференцировке. Во взрослом организме эти агенты участвуют в контроле клеточного цикла, а i-cdk останавливают пролиферацию нейронов, предотвращая появление опухолей.
Выход клетки из покоящегося состояния (фазы G0) и запуск фазы G1 обеспечивается так: митогенные стимулы активируют митоген-зависимые протеинкиназы (МАРК) и экспрессию циклина D (CnD) (рис. 1):
|
|
|
|
|
|
|
|
Рис. 1. Схема клеточного цикла.
Далее происходит образование комплекса CnD/cdk4 (или cdk6), который обеспечивает переход клетки из фазы G0 в фазу G1. Если в этот период экспрессия CnD подавляется (воздействием МАРК, активированных, например, в результате клеточного стресса), то происходит остановка клеточного цикла на стадии G1. В начале фазы G1 в клетке появляется CnE (деактивация которого также приводит к остановке клеточного цикла в G1 фазе). Для осуществления перехода в следующую фазу цикла – S фазу, необходимо образование CnE/cdk2 комплекса. В этой же G1 фазе начинает экспрессироваться CnA; и комплекс CnA/cdk2 обеспечивает вступление клетки в следующую (после стадии S) фазу G2 клеточного цикла. Недостаток CnA влечет за собой остановку клеточного цикла в G2. CnB, присутствующий в клетке в течение всей интерфазы, активируется в самом конце фазы G2 и образует комплекс с cdс2, необходимый для начала последней стадии цикла - М фазы. Деактивация CnB приводит к остановке клеточного цикла в фазе G2 [ 12, 33 ].
1.3 Ишемический инсульт и нейрогенез
Ишемический инсульт (инфаркт мозга) развивается при значительном уменьшении мозгового кровотока. На стадии реперфузии, когда мозговой кровоток восстанавливается, происходит интенсификация окислительного стресса, который характеризуется как дисбаланс между продукцией активных форм кислорода и емкостью антиоксидантной защиты. В результате окислительного стресса происходит повреждение липидов, белков и нуклеиновых кислот и гибель клеток мозга по некротическому и/или апоптотическому типу.
Гибернация является уникальным физиологичеким состоянием, при котором происходит снижение метаболизма и температуры тела, обеспечивающие выживание в период дефицита пищи. Значительная степень нейропротекции при гибернации обусловливается комбинацией факторов, играющих важную роль в защите мозга от повреждения при кардинальном изменении мозгового кровотока во время спячки и после пробуждения. Адгезия нейтрофилов и инфильтрация макрофагов в место повреждения стимулируют цитотоксичекие реакции при ишемии мозга. Лейкоцитопения, снижение продукции антител и повышение антиоксидантной защиты могут защищать нейроны от повреждения при гибернации. На моделях фокальной ишемии-реперфузии показано, что ингибирование синтеза белка, которое также наблюдается при гибернации, обладает нейропротекторным действием [ 22 ].
Известно, что в мозге экспериментальных животных после ишемии усиливается нейрогенез, который частично компенсирует нейродегенеративные изменения. Показано, что в моделях на грызунах инсульт, вызванный окклюзией средней мозговой артерии (ОСМА), запускает усиленный нейрогенез в поврежденном стриатуме и неповрежденном гиппокампе молодых грызунов. В результате экспериментов на животных старшего возраста было показано, что у молодых и старых крыс происходит одинаковое увеличение числа новообразованных нейронов стриатума после инсульта, несмотря на то что базальная пролиферация клеток в субвентрикулярной зоне у старых крыс была снижена [2]. Напротив, число новообразованных после инсульта гранулярных клеток и базальный нейрогенез в субгранулярной зоне зубчатой извилины были ниже у старых животных по сравнению с молодыми. Кроме того, у старых крыс в зубчатой извилине была ослаблена способность новообразованных клеток дифференцироваться в нейроны. Но, тем не менее, величина стриарного постинсультного нейрогенеза сопоставима у молодых и старых животных, что указывает на функционирование этого механизма восстановления также и в пожилом возрасте.
Таким образом, механизмы нейропротекции при гибернации связаны с гипотермией, лейкоцитопенией, снижением синтеза белка, повышением антиоксидантного статуса клеток мозга и , по-видимому, могут включать в себя также и изменение пролиферации клеток мозга ни только в направлении образования новых нейронов, как показано на моделях ишемического повреждения мозга, но и в пролиферации эндотелиальных клеток сосудов мозга и клеток глии [ 24 ].
Глава 2. Материалы и методы
2.1 Объект исследования
В работе использовали отделы мозга взрослых сусликов Citellus undulatus обоего пола массой 280-400 г, отловленных в Якутии.
2.2 Условия гибернации
Животных содержали в индивидуальных ячейках, снабжённых термодатчиком, в условиях естественного освещения при свободном доступе к воде и пище. В середине октября сусликов переносили в специальное помещение, где они впадали в спячку при температуре воздуха 4°С и находились там до полного пробуждения (середина апреля). Опыты проведены на 7 группах сусликов, взятых в разные фазы гибернационного цикла и на разных стадиях снижения и повышения температуры тела:
1 группа (n=8) – бодрствующие активные в летний период (июнь) животные, контроль;
2 группа (n=7) – животные, входящие в спячку осенью при температуре тела +36 ˚С ;
3 группа (n=7) – животные, входящие в спячку осенью при температуре тела +10 ˚С ;
4 группа (n=10) – спящие животные (январь),середина баута;
5 группа (n=4) – спонтанно пробудившиеся животные, межбаутное бодрствование;
6 группа (n=10) – животные, пробудившиеся весной при температуре тела +25 ˚С;
7 группа (n=5) – животные, пробудившиеся весной при температуре тела +10 ˚С.
Моделирование состояния зимней спячки у сусликов осуществляли сотрудники института Биофизики клетки РАН (Пущино-на-Оке).
2.3 Подготовка образцов тканей мозга для анализа
Мозг сусликов вынимали, охлаждали в ледяном изотоническом растворе NaСl и на льду выделяли кору больших полушарий, мозжечок, гиппокамп, гипоталамус и ствол мозга. Образцы замораживались и хранились в жидком азоте, а затем были любезно предоставлены нам для анализа ведущим научным сотрудником института Биофизики клетки РАН (Пущино-на-Оке) д.б.н. Семеновой Т.П.
Ткани гомогенизировали при 4°С в гомогенизаторе Поттера (1500 об/мин) в течение 15-20 сек в 5 объемах буфера (HEPES 20мМ рН 7,4). Затем гомогенаты центрифугировали 30 мин при 13000 g и 4°С. Полученный супернатант аликвотировали, замораживали и использовали в дальнейших исследованиях.
2.4 Биохимические методы исследования
2.4.1 Определение содержания белка
Концентрацию белка определяли по методу Бредфорд [ 16 ]. К 10 мкл супернатанта в оптимальном разведении добавляли 500 мкл 0.01% раствора красителя Кумасси ярко синего G-250 и инкубировали 15 мин при комнатной температуре. После этого измеряли поглощение при 595 нм. В качестве калибровочного стандарта использовали бычий сывороточный альбумин в диапазоне концентраций 0,1-0,8 мг/мл. В этом диапазоне концентраций стандартного белка поглощение при 595 нм линейно зависит от концентрации белка и поэтому все пробы, после предварительного эксперимента, разводили до получения оптической плотности, соответствующей линейному участку у стандарта.
2.4.2 Электрофорез в полиакриламидном геле. Вестерн-блот
Электрофорез в ПАГГ применяется для разделения белков по молекулярной массе под действием электрического тока. В качестве стандартного образца используют белки-маркеры с молекулярными массами 11-170 кДа.
Для разделения исследуемых белков использовался 10% разделяющий (нижний) гель (2,5 мл 30% АА, 0,8% bisАА; 1,875 мл 4хTris/Cl/SDS pH=8,8; 3,125 мл Н2О; 25 мкл 10% аммония персульфата; 5 мкл TEMED) и 3,9% концентрирующий (верхний) гель (0,65 мл 30% АА, 0,8% bisАА; 1,25 мл 4хTris/Cl/SDS pH=6,8; 3,05 мл Н2О; 25 мкл 10% аммония персульфата; 5 мкл TEMED) .
Подготовка проб к электрофорезу осуществлялась следующим образом: к аликвоте супернатанта объёмом 100 мкл добавляли 20 мкл Sample buffer, кипятили 5 мин при 100°С и охлаждали до комнатной температуры. Затем пробы центрифугировали 5 мин при 13000 g и комнатной температуре и отбирали супернатант, который перед нанесением на гель разводили таким образом, чтобы конечная концентрация белка в нём составляла 60 мкг на лунку геля.
Электрофоретическое разделение исследуемых белков проводилось в денатурирующих условиях (использовался 1х Running buffer, содержащий 1% додецилсульфата натрия (Tris, глицин, SDS)) 1-1,5ч при 200 V. Перенос белков с геля на мембрану PVDF осуществлялся в Transfer buffer (Tris, глицин,этиловый спирт) 45-60 мин при 150 mА,110 V.
2.4.3 Выявление белков клеточного цикла и циклинзависимой киназы Cdk5
Мембрану с перенесенными белками блокировали в течение 15 – 16 ч для предотвращения неспецифического связывания в 5% растворе обезжиренного молока в TBST (50 мМ Tris/Cl , pH=7,5; 0,9% NaCl; 0,05% Tween 20), после чего инкубировали с различными разведениями специфических первичных антител в 5% молоке в TBST 2ч при комнатной температуре или 15-16ч при 40С. Для удаления несвязавшихся антител мембрану отмывали 4-х кратно в TBST в течение 30 мин. Связавшиеся первичные антитела выявляли с помощью вторичных антител, полученных у другого вида животных против первичных, конъюгированных с пероксидазой хрена. Связывание антител визуализировали на рентгеновской пленке с использованием хемилюминесцентной системы, которая в присутствии пероксидазы хрена окисляется с выделением квантов света.
Детекцию белков проводили в оптимальных условиях, которые были подобраны ранее.
Таблица 1
Условия иммунодетекции исследуемых белков при Вестерн-блот анализе.
| Белок | 1 Ab | Разведение | 2Ab | Разведение |
| MCM 2 | Goat polyclonal (козьи поликлональные) | 1:500 | Rabbit-anti-Goat (кроличьи антикозьи) | 1:2000 |
| Cn B1 | Rabbit polyclonal (кроличьи поликлональные) | 1:1000 | Goat-anti-Rabbit (козьи антикроличьи) | 1:2000 |
| Cn A | Rabbit polyclonal | 1:1000 | Goat-anti-Rabbit | 1:1000 |
| Cdc 2 | Rabbit polyclonal | 1:1000 | Goat-anti-Rabbit | 1:2000 |
| Cdk 2 | Rabbit polyclonal | 1:500 | Goat-anti-Rabbit | 1:2000 |
| Cdk 4 | Rabbit polyclonal | 1:1000 | Goat-anti-Rabbit | 1:2000 |
| Cdk 5 | Rabbit polyclonal | 1:1000 | Goat-anti-Rabbit | 1:2000 |
2.5 Статистическая обработка результатов
Статистический анализ осуществлялся с помощью программы Statistica 7 с использованием t-критерия Стьюдента.
Различие считалось достоверным при уровне значимости Р<0,05.
Все исследования выполнены в лаборатории функциональной биохимии нервной системы института Высшей нервной деятельности и нейрофизиологии РАН.
Автор выражает глубокую благодарность за оказанную всестороннюю помощь при выполнении работы сотрудникам лаборатории функциональной биохимии нервной системы института Высшей нервной деятельности и нейрофизиологии РАН, отдельно к.б.н. Онуфриеву Н.В. и д.б.н., профессору Гуляевой Н.В.
Глава 3. Полученные результаты и их обсуждение
Экспрессия белков клеточного цикла и Cdk 5 имеет различный уровень в исследованных отделах мозга и зависит от функционального состояния сусликов.
Гибернация достоверно снижает в гиппокампе уровень экспрессии Cdc 2, Cdk 2 и Cdk 4, а экспрессия MCM 2 и Cdk 5 имеет лишь тенденции к снижению по сравнению с контролем (активные животные) (рис.2) . Экспрессия циклинов. А и В1 остаётся практически на том же уровне, что и у активных сусликов. У спонтанно пробудившихся животных уровень экспрессии исследуемых белков также существенно не изменялся по сравнению с контролем. Однако на стадии пробуждения при температуре + 25°С была выявлена положительная корреляция между циклином В1 и МСМ 2 (r = 0,97 ,p < 0,05). Возможно, на стадии пробуждения при температуре + 25°С образование МСМ 2 идет однонаправленно с биосинтезом Сn B1 в отличие от пробуждения при температуре + 10°С (известно, что комплекс МСМ 2/МСМ4 является субстратом для комплекса Cdc2/Cn B1 in vitro). МСМ 2 является маркёром репликации ДНК, его уровень возрастает при переходе клетки из фазы G1 в S фазу клеточного цикла. А циклин В1 активируется в конце фазы G2, образуя комплекс с Cdc 2, небходимый для начала последней стадии цикла – М фазы (собственно митоза).
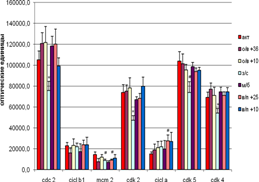
Рис. 2 Экспрессия исследуемых белков в гиппокампе сусликов CITELLUS UNDULATUS на разных стадиях гибернационного цикла
Отличия от группы контрольных животных: * - p < 0,05; # - p < 0,1.
В коре больших полушарий мозга на экспрессию белков влияла температура окружающей среды, при которой животные находились на стадиях вхождения в спячку и выхода из неё (рис. 3). Достоверными являются различия уровня экспрессии Cdk 5 и Cdk 4 внутри групп вхождения и выхода из спячки; Cdc 2 и MCM 2 – внутри группы входа из спячки; циклина В1 – внутри группы вхождения в спячку. Так,по сравнению с контрольной группой животных на стадии пробуждения при +10°С достоверно возрастал уровень экспрессии Cdc 2, MCM 2 и Cdk 4 . При +25°С достоверно снижалась экспрессия Cdk 4; на уровне тенденций экспрессия Cdc 2 снижалась, а циклина В1 и MCM 2 - повышалась. При вхождении в спячку при +10°С достоверно возрастала экспрессия MCM 2 и Cdk 4,снижалась экспрессия Cdk 5 ; а при + 36°С на уровне тенденций изменялась только экспрессия Cdk 4 (уменьшалась).
У спящих животных по сравнению с контрольной группой различий в экспрессии исследуемых белков не наблюдалось. В межбаутный период достоверно уменьшалась экспрессия циклина В1 и увеличивалась экспрессия Cdk 4, экспрессия Cdk 5 имела тенденцию к снижению. В межбаутный период была выявлена отрицательная корреляция между Cdk 2 и циклином В1 (r = - 0,98 ,p < 0,05).
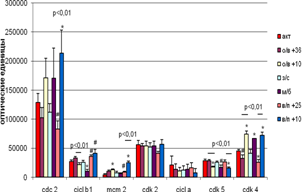
![]()
Рис. 3 Экспрессия исследуемых белков в коре больших полушарий мозга сусликов CITELLUS UNDULATUS на разных стадиях гибернационного цикла
Отличия от группы контрольных животных: * - p < 0,05; # - p < 0,1.
В мозжечке не наблюдается достоверных различий внутри групп вхождения и выхода из спячки при различных температурных режимах содержания животных (рис. 4). Однако на стадии вхождения в спячку (в сравнении с контролем) наблюдается достоверное увеличение экспрессии Cdk 2 как при + 36°С, так и при + 10°С, а Cdk 4 – только при + 10°С. Экспрессия МСМ2 и циклина А увеличивается на уровне тенденций. Выход из спячки при + 10°С сопровождается достоверным увеличением экспрессии Cdc 2 и Cdk 2 , а Cdk 4 и Cdk 5- увеличением на уровне тенденций. При пробуждении при температуре +25°С тенденции к увеличению экспрессии имеют циклин В1, МСМ2, Cdk 2 , Cdk 4 и Cdk 5.
У спящих животных достоверно снижалась экспрессия только Cdc 2.
Успонтанно пробудившихся животных достоверно возрастала экспрессия только Cdk 4, а количество остальных белков колебалось на уровне контроля.
Положительная корреляция была выявлена между Cdc 2 и циклином В1 (r = 0,97 ,p < 0,05) на стадии вхождения в спячку при температуре + 10°С , отрицательная – на стадии пробуждения при температуре + 25°С между Cdk 2 и циклином А. Наверное различную роль играют Cdc 2 и циклин В1 на стадии вхождения в спячку, когда пролиферация снижается и при пробуждении, когда она более вероятна.
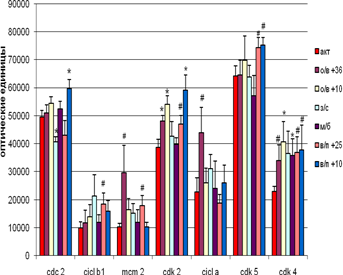
Рис.4 Экспрессия исследуемых белков мозжечке сусликов CITELLUS UNDULATUS на разных стадиях гибернационного цикла
Отличия от группы контрольных животных: * - p < 0,05; # - p < 0,1.
В стволе мозга гибернация не вызывала достоверных изменений в экспрессии исследуемых белков, только уровень МСМ2 имел тенденцию к увеличению.
В межбаутный период экспрессия белков изменялась недостоверно.
Уровень экспрессии циклина А достоверно повышался при вхождении в спячку при температуре +36˚С и при выходе из неё при температуре +25˚С, на уровне тенденции – при пробуждении при температуре +10˚С. Достоверным является различие в экспрессии Cdk 2 в группе выхода из спячки при разных температурных режимах (при температуре +25˚С – уменьшалась, а при +10˚С - увеличивалась ). Различия наблюдаются только внутри группы и отсутствуют при сравнении с активными и межбаутными. Хотя при низкой температуре тела во время выхода выхода из спячки экспрессия некоторых белков в основном повышается!
Отрицательные корреляции были выявлены между Cdk 2 и циклином А (r = -0,99 ,p < 0,05) на стадии пробуждения при температуре +10˚С и между Cdk 2 и циклином В1 при вхождении в спячку при температуре +36˚С.
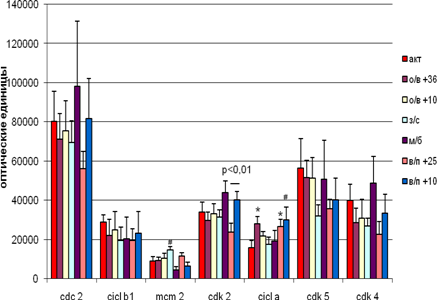
Рис. 5 Экспрессия исследуемых белков стволе мозга сусликов CITELLUS UNDULATUS на разных стадиях гибернационного цикла
Отличия от группы контрольных животных: * - p < 0,05; # - p < 0,1.
Особенностью метаболизма мозга является интенсивный окислительный обмен. Основным субстратом энергетического метаболизма мозга является глюкоза [ 14 ]. Клетки головного мозга утилизируют 20% кислорода, потребляемого организмом, хотя сам мозг составляет около 2% от массы тела [ 14, 17 ], что обуславливает потенциальную возможность образования большого количества АФК в процессе окислительного фосфорилирования. Это обстоятельство предопределяет потребность мозга в высоком уровне кровоснабжения.
В различные периоды гибернационного цикла существенно меняется интенсивность метаболических процессов и интенсивность энергетического обмена в тканях, в том числе и головном мозге [ 35, 39 ]. Но именно ведущая роль в функциональных нейрохимических процессах в мозге принадлежит несомненно белково – нуклеиновому метаболизму [ 3 ]. В течение спячки сусликов наблюдается подавление синтеза белков в головном мозге и очень резкая активация образования белков при побуждении [ 4 ]. Поэтому можно ожидать, что уровень экспрессии белков клеточного цикла и Сdk 5 в различных отделах мозга будет зависеть от физиологического состояния животных в период гибернации.
Ряд данных указывает на то, что при гибернации повышается устойчивость головного мозга к повреждающим факторам. Например, Фрерих и Халенбек установили, что срезы гиппокампа у гибернирующих 30-ти линейных сусликов имеют повышенную толерантность к гипоксии и агликемии [ 25 ]. Зои с сотр. показали, что в состоянии зимней спячки значительно снижается в мозге вызванный повреждением окислительный стресс [ 42].
При гибернации существенно изменяется липидный состав мембран клеток мозга [ 7 ]. Кроме того, при переходе сусликов от бодрствования к спячке происходит латеральное разделение липидов мембран [ 15 ]. В результате этого образуются большие липидные области содержания ненасыщенных жирных кислот, а белки скапливаются в отдельные узкие зоны. Изменение липидного состава и их упаковки при гибернации направлено на сохранение структурной целостности и функциональных свойств мембран.
Последние данные показывают, что в мозге гибернирующих сусликов связанный с микротубулином белок tau гиперфосфорилирован по всем шести сайтам (S199, T205, S214, S262, S396, S404). Интересно, что только три из этих сайтов (S199, S262 и S404) являются дефосфорилированными у пробуждающихся животных, что предполагает обратимое фосфорилирование указанных сайтов [ 28 ]. У активных летом сусликов наблюдается самый низкий уровень уровень фосфорилирования. Известно, что белок tau аномально гиперфосфорилирован в мозге особей с болезнью Альцгеймера и других tau – патологий и, как предполагают, играет важную роль в патогенезе этих заболеваний. А поскольку механизмы, приводящие к аномальному фосфорилированию tau остаются неясными, то последние наблюдения обратимого фосфорилирования этого белка во время гибернации являются идеальной физиологической моделью для изучения данного процесса in vivo.
Выводы
1. Экспрессия белков клеточного цикла и Cdk 5 имеет различный уровень в исследованных отделах мозга и зависит от функционального состояния сусликов.
2. Температура тела сусликов влияет на экспрессию белков клеточного цикла и Cdk 5 в коре больших полушарий и стволе мозга на стадиях вхождения в спячку и выхода из неё.
3. Наличие корреляционных связей между циклинами и соответствующими киназами позволяет предположить, что в отделах мозга сусликов на стадиях вхождения в спячку и выхода из неё появляются клетки на поздних фазах клеточного цикла.
Литература
1. Белоусов А.Б. Роль центральной нервной системы в контроле зимней спячки // Усп. физиол. наук. – 1993. – Т. 24, № 2. – С. 109-126.
2. Дарзалиа В., Хелманн У., Линдвалл О., Кокаиа З. Нейрогенез в стареющем головном мозге после инсульта // Инсульт. – 2007, № 1.
3. Демин М.М., Шортанова Т.Х., Эмирбеков Э.З. Нейрохимия зимней спячки млекопитающих. Л.: Наука, 1988. –137 с.
4. Жегунов Г.Ф. , Микулинский Ю.Е. Активация синтеза белка в тканях сусликов при пробуждении после зимней спячки // Укр. биохим. журн. 1987. Т. 59, № 3. С. 69-73.
5. Игнатьев Д.А., Сухова Г.С. Сухов В.П. Анализ изменений частоты сердцебиений и температуры суслика Citellus Undulatus в различных физиологических состояниях // Общая биология – 2001. – Т. 62, №1. – С. 66-77.
6. Калабухов Н.И. Спячка млекопитающих. М.: Наука, 1985. – 259 с.
7. Коломийцева И.К., Перепелкина Н.И., Патрушев И.В., Попов В.И. Роль липидов в сборке эндоплазматического ретикулума и диктиосом якутского суслика Сitellus undulates при гибернации // Биохимия. – 2003. – Т. 68, вып. 7. – С. 954-967.
8. Крепс Е.М. Липиды клеточных мембран. Л.: Наука, 1981. - С.339.
9. Соломонов Н.Р. Основные итоги и задачи изучения зимней спячки грызунов Якутии // Эколого-физиологические характеристики природных гипометаболических состояний. – Пущино, 1992. – С. 29-34.
10. Штарк М.В. Мозг зимоспящих. Новосибирск: «Наука», 1970. – 240 c.
11. Шугалей Л.С. Молекулярные основы устойчивости зимоспящих животных к неблагоприятным условиям среды // Эколого-физиологические характеристики природных гипометаболических состояний: Сб. науч. тр. / Под ред. Колаевой С.Г. – Пущино, 1992. – С. 70-73.
12. Эмирбеков Э.З., Львова С.П., Кличханов Н.К. Биохимические изменения в крови при искусственной и естественной гипотермии // Пробл. криобиол. – 1995. - №1. – С.14-21.
13. Эмирбеков Э.З., Львова С.П., Мусаев Б.С., Мейланов И.С., Симмалавонг Сантисук, Бутаева П.Ш., Абдулаева М.З. Роль липидов мозга при гипотермии и самосогревании гомойотермных и зимоспящих животных // Биохим. аспекты холодовых адаптаций: Сб. науч. тр. - Харьков, 1991. – С. 166-175.
14. Ames I.A. CNS energy metabolism as related to function // Brain Research Rew. – 2000. – V. 34. – P. 42-68.
15. Azzam N.A., Hallenbeck J.M., Kachar B. Membrane changes during hibernation // Nature. – 2000. – V. 407. – P. 317-318.
16. Bradford M.M. A rapid and sensitive method for quantitation of microgram quantities of protein using the principle of protein bindong // Anal. Biochem. 1976. – V. 72. – P. 248-254.
17. Clarke D.D., Sokoloff L. Circulation and energy metabolism of the brain // Basic Neurochemistry: Molecular, Cellular and Meducal Aspects. Sigel G.J., Agranoff B.W., Alberts R.W., Fisher S.K., Uhler M.D. (Eds.) – Phyladelphia:Lippincott – Raven. – 1999. – P 637-669.
18. Christina G. von der Ohe, Craig C. Garner, Corinna Darian-Smith, H. Craig Heller. Synaptic Protein Dynamics in Hibernation. // Neurosciens. – 2007.- Nov 9; 149 (3).
19. Christina G. von der Ohe, Craig C. Garner, Corinna Darian-Smith, H. Craig Heller. Ubiquitos and Temperature - Dependent. // Neurosciens. – 2008.- Apr 9; 152 (4).
20. Copani Agata; Uberti Daniela; Sortino Maria Angela; Bruno Valeria; Nicoletti Ferdinando and Memo Mauritio. Activation of cell-cycle-associated proteins in neuronal death: a mandatory or dispensable path? Trends in Neuroscience. 2001 Jan; 24 (1): 25-31.
21. Copani А.; Sortino M. A.; Nicoletti F. and Stella Giuffrida A. M. Alzheimer’s Disease Research Enters a “New Cycle”: How Significant? // Neurochemical Research. 2002 Feb; 27 (Nos. 1/2): 173-176.
22. Drew K.L., Rice M.E., Kuhn T.B., Smith M.A. Neuroprotective adaptations in hibernation: therapentic implications for ischemia-reperfusion, traumatic brain injury and neurodegenerativedislases // Free Radic. Biol. Med. – 2001. –V. 31. – P. 563-573.
23. Droge W. Free Radicals in the Physiological control of cell Function // Physiol. Rev. – 2002. – V. 82. – P. 47-95.
24. Fang Zhou, Xiongwei Zhu, Rudy J. Castellani,Raphaela Stimmelmayr, George Perry, Mark A. Smith, and Kelly L. // Drew Hibernation, a Model of Neuroprotection // American Journal of Pathology, V. 158, No. 6, June 2001.
25. Frerichs K.U., Kennedy C., Sokoloff L., Hallenbeck J.M. Local cerebral blood flow during hibernation, a model of natural tolerance to «cerebral ishemia» // J Cereb Blood Flow Metab. – 1996. – V. 14. – P. 193-205.
26. Frerichs K.U., Hallenbeck J.M. Hibernation in ground squirrels induces stats and species-specific tolerance to hypoxia and aglycemia: an in vitro study in hippocampal slices // J. Cereb. Blood Flow. Metab. – 1998. – V. 18. – P. 168-175.
27. Hermes-Lima M., Zentenc-Sakin T. Animal response to drastic changes in oxygen akailability and physiological oxidative stress // Comp. Biochem. Physiol. C. Toxicol. Pharmacol. – 2002. – V/ 133/ - P. 537-556.
28. Jens Thorsten Stieler, Torsten Bullman, Franziska Kohl, Brian M. Barnes, Thomas Arendt. Physiological regulation of tau phosphorylation during hibernation. // J Neural Transm. – 2009 (116): 345 – 350.
29. Lipton Р. Ischemic cell death in brain neurons // Physiol. Rev. – 1999. – V. 79. – P. 1431-1568.
30. Nagy Zs. Cell cycle regulatory failure in neurones: causes and concequences. Neurobiology of Aging. 2002 (21): 761-769.
31. Nagy Zs.; Esiri M. M.; Cato A.-M. and Smith A. D. Cell cycle markers in the hippocampus in Alzheimer's disease // Acta Neuropathol. 1997 (94): 6-15.
32. Nurnberger F. The neuroendocrine system in hibernating mammals // Cell and Tissue. Res. – 1995. – V. 28. – P. 391-412.
33. Ohnuma Shin-ichi; Philpott Anna and A Harris William. Cell cycle and cell fate in the nervous system. Current Opinion in Neurobiology. 2001 (11): 66-73.
34. Storey K.B. Turning down the fires of life: metabolic regulation of hibernation and estivation // Mol. mech. metab. Arrest. – 2001. – P. 1-21.
35. Storey K.B., Storey J.M. Metabolic rate depression in animals: transcriptional and translational controls // Biol. Rev. Camp. Philos. Soc. – 2004. – V. 79. – P. 207-233.
36. Toien O., Drew K.L., Chao M.L., Rice M.E. Ascorbat dynamics ana oxygen consumption during arousal from hibernation in arctic ground squirrels // Am. J. Physiol. – 2001. – V. 281. – P. 572-583.
37. Van Brenkelen F., Martin S.L. Reversible depression of transcription during hibernation // J. Comp. Physiol. (B). – 2002-V. 172. – P. 355-361.
38. Wang L.C.H. Energetic and field aspects of mammalian torpor: the Richardson groundsquirrel // Strategus in Cold: Natural Torpidity and Thermogenesis // Eds L.C.H. Wang, J.W. Hudson, N.Y.: Acad. Press. – 1978. – P. 109-145.
39. Wang L.C.H., Lee T.F. Torpor and hibernation in mammals: metabolic, physiological, and biochemical adaptations // Handbook of physiology: Oxford univer. Press., 1996. - № 4. – P. 507-531.
40. Yeh J., Tam C.F., Catuirs E., Le T.T., Papa V., Pena L., Vasquez M., Vu C., Wang S., Lopez G.A. Changes in karious plasms lipid components, glucose, and insulin in spermophilus lateralis during hibernation // Comp. Biochem. Physiol. B. Biochem. Mol. Biol. – 1996. – V. 111. – P. 651-663.
41. Young J.S., Woodside J.V. Antioxidants in health and disease // J. Clin. Pathol. – 2001. V. 54. – P. 176-186.
42. Zhou G., Zhu X., Castellani R.J., Stinnelmayr R., Perry G., Smith M.A., Drew K. Hibernation, a model of neuroprotection // Am. J. Pathol. – 2001. – V. 158. – P. 2145-2151.