Курсовая работа: Сезонная динамика фитопланктона Средней Оби в 2007-2008 гг.
Федеральное агентство по образованию
ТОМСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ (ТГУ)
Биологический институт
Кафедра ихтиологии и гидробиологии
ВЫПУСКНАЯ РАБОТА
СЕЗОННАЯ ДИНАМИКА ФИТОПЛАНКТОНА СРЕДНЕЙ
ОБИ В 2007-2008 ГОДУ
Ильякова Анастасия Михайловна
Руководитель
н. с. НИИББ
О.С. Голубых
Автор работы
Студентка ІV курса
А.М. Ильякова
2009г
Оглавление
Введение
1. Материал. Методика сбора
2. Физико-географическая характеристика исследуемого водоема
3. Ценотическая характеристика групп водорослей в 2008 году
4. Характеристика биотопов Средней Оби по микроводорослям в 2008г
5. Сезонное развитие фитопланктона Средней Оби в 2008 г.
6. Определение сапробности исследуемого участка реки Обь в 2008 году
7. Сравнение динамики развития фитопланктона за 2007-2008 года
Заключение
Список используемой литературы
Приложения
Введение
Огромное значение в экономическом развитии Сибири имеют реки. Они являются носителями гидроэнергии, используются для развития рыбного хозяйства, служат источником водоснабжения. Обострение экологической обстановки на реках требует проведение их мониторинга. В комплексных разработках научных основ управления водными экосистемами видное место занимают исследования фитопланктона, как первого звена трофической цепи, которое во многом определяет структуру и функционирование водной экосистемы в целом. Фитопланктон Оби играет значительную роль в создании первичного органического вещества и как фактор формирования качества воды. Фитопланктон чутко реагирует на изменения экологических условий, его продукция определяет трофический уровень водоема, а состав и обилие характеризует его санитарное состояние. Как известно, водоросли выступают в качестве биологического индикатора и способны фиксировать незначительные изменения в экосистеме, не обнаруживаемые другими методами исследований. Все это определяет повышенный интерес к исследованию данной группы гидробионтов. Сибирь в альгологическом отношении изучена недостаточно. Это касается и одной из крупнейших рек планеты - Оби. По своим природным условиям Обь неоднородна. Крупные ссоры, обилие перекатов, множество притоков, особенно в среднем и нижнем течении, различаются по особенностям состава планктонных водорослей, уровню развития, пространственному распределению, специфике сезонной и межгодовой динамики. Для познания вопросов, связанных с функционированием экосистемы Оби во временном аспекте, крайне важно знать, как работает ее первичное звено - фитопланктон. Знание видового состава растительного планктона, их количественного изменения по сезонам необходимо для оценки качества вод реки при использовании ее в водоснабжении и рыбном хозяйстве.
Цель работы - альгофлористическая и временная характеристики фитопланктона Средней Оби, сравнение динамики микроводорослей за 2007-2008 года и оценка сапробиологического состояния реки.
Для решения этих вопросов были поставлены следующие задачи: выяснение состава фитопланктона, сезонной динамики численности, изучение биологии доминирующих видов, оценка состояния вод Оби на основе выявления видов-эдификаторов и редких видов водорослей.
1. Материал. Методика сбора
Материалом для написания данной работы послужили пробы, собранные у п. Половинка Томского района. В период летней практики было поставлено три опыта на определение продукции фитопланктона, взято 12 проб для определения содержания пигментов в воде и 18 проб на видовое разнообразие фитопланктона.
Качественные пробы объемом 0,5л брали 2 раза в месяц на протяжении 7 месяцев с апреля по октябрь 2008 года у правого берега реки, на границе верхней и средней Оби, выше впадения в нее р. Томь. Тип грунта на данном участке галечно-песчано-глинистый; берег достаточно пологий. Растительности мало, растет небольшими скоплениями осока. и тальники.
Пробы отбирались с поверхности водоема в бутылки по 0,5л и фиксировались нейтрализованным формалином до появления слабого запаха.
В лаборатории пробы концентрировались до 50 мл в воронках сгущения Сорокина на фильтрах нуклеопор с диаметром отверстий 1-2мкм.
Качественная обработка фитопланктона проводилась с использованием микроскопа МБИ-11, а также общепринятых в альгологии определителей и руководств. Пробы просматривались на увеличении 10*40. При этом сразу делались карточки с описанием каждого вида, его местообитания, экологии.
Количественная оценка фитопланктона проводилась визуально. Для просчета использовалась камера Нажотта, представляющая собой пластинку из толстого стекла толщиной до 0,5см, ширина и толщина которой соответственно 2,5*6,5см. В центральной ее части располагается круглое углубление, в котором помещена решетка, разделенная на 48 прямоугольных отделов. В это углубление помещается пипеткой капля из пробы фитопланктона. Длина, ширина и глубина камеры над решеткой составляет 7,9мм, 6,1мм и 0,88мм соответственно. Следовательно, объем равен 42,4072мм³.
Так как просмотреть все 48 полосок очень затруднительно по причине больших временных затрат, а также может высохнуть капля воды с пробой, поэтому обычно просчитывают лишь часть камеры (например,6,8 или 12 полос, домножая соответственно на 8, 6 и 4). Таким образом, получают достаточно усредненные данные. Для большей точности просчет одной пробы проводят, по меньшей мере, 3 раза, каждый раз беря пипеткой новую порцию пробы. Затем вычисляют среднюю арифметическую.
Для определения продукции фитопланктона в данной работе использовали метод измерения скорости фотосинтеза в воде, заключенной в склянки, в его кислородной модификации, предложенной Г.Г. Винбергом в 1934г.
В работе используются склянки из белого и темного стекла с притертыми пробками и с точно известным объемом каждой. В постановке опыта использовалось 7 склянок. Три склянки использовались в качестве темных, они помещались в черные мешочки, три в качестве светлых и одна - контрольная. Склянки при заполнении должны быть погружены в воду, чтобы исключить попадание в них пузырьков воздуха. Время экспозиции склянок составляет 24 часа. После этого склянки снимаются с установки и в них сразу фиксируется кислород. Для фиксации приливается по 1мл растворов хлорида Mn (32%) и KI+NaOH (растворы KI (10%) и NaOH (32%) сливаются в объемах 1:
1). Затем склянки доставляются в лабораторию для дальнейшей обработки. Определение содержания в воде кислорода основано на измерении количества образующегося осадка из йода, учитывающегося титрованием раствора тиосульфата. Осадок должен отстояться не менее 10 минут, затем в склянку добавляется 5мл раствора серной кислоты для его растворения. Склянка закрывается пробкой, и содержимое тщательно перемешивается. После производится его титрование до светло-желтого цвета. Далее прибавляется 1мл свежеприготовленного раствора крахмала (0,1%) и продолжается титрование до исчезновения синей окраски. Объем ушедшего на титрование тиосульфата записывают и затем используют для расчета кислорода.
Далее ведется расчет продукции фитопланктона. Валовую первичную продукцию за время экспозиции склянок получают по разности содержания кислорода в светлой и затемненной склянках к концу их экспозиции в реке.
По убыли содержания растворенного кислорода в затемненной склянке, по сравнению с исходной его концентрацией, судят о скорости деструкции органического вещества, эквивалентно связанной с потреблением кислорода планктонным сообществом.
Разность между валовой продукцией и деструкцией дает чистую первичную продукцию планктона в целом.
2. Физико-географическая характеристика исследуемого водоема
Река Обь - главная водная артерия Западной Сибири. Она образуется при слиянии рек Бии и Катуни, берущих начало на Алтае, и впадает в обскую губу Карского моря.
Рельеф области, по которой протекает река, отличается исключительной равнинностью и сравнительно слабой расчлененностью. Максимальные абсолютные отметки поверхности (до 258м) находятся на юго-востоке области, приуроченные к северным отрогам Кузнецкого Алатау. Отсюда поверхность понижается в северо-западном направлении до 40-80м над уровнем моря. В этом же направлении протекает и река Обь, которая делит область на две почти равные части: более возвышенное правобережье и левобережье, на юге которого расположено крупнейшее в мире Васюганское болото с абсолютными отметками, достигающими 166м. Склоны междуречий очень пологие (0-1˚), лишь у речных долин крутизна их увеличивается до 30˚ (Н.С. Евсеева, А.А. Земцов, 1990).
Обь занимает по протяженности 7 место в Евразии и 16 место в мире, по площади водосбора - соответственно 1 и 6 места. Ее протяженность 3676км, средний годовой сток 394км³, водосборная площадь - 2990км².
Обь имеет самые низкие уклоны из всех крупных рек, впадающих в Северный ледовитый океан (Малик Л.К., 1978). Средний уклон Оби по территории Томской области, как правило, лежит в пределах 0,03-0,04м/км. Обь представляет собой типично равнинную реку с малым падением, обширной долиной и поймой. По условиям формирования речного стока река преимущественно снегового питания: доля снегового питания Оби составляет в среднем 49%, дождевого - 27%, грунтового - 16%, ледникового - 8%. (А.М. Догановский, 2004)
Тип климата региона определяется как переходный от умеренно-континентального Восточно-Европейской равнины к резко-континентальному климату Восточной Сибири. Над данной территорией, как летом, так и зимой преобладают континентальные умеренные воздушные массы, что ведет к увеличению температуры летом и снижению ее зимой. Территория области отличается суровой продолжительной зимой с сильными ветрами, устойчивым снежным покровом и довольно жарким летом. Переходные периоды короткие, с резкими колебаниями температуры. Наиболее низкие среднемесячные температуры в регионе наблюдаются в январе (до - 21,5˚С), наиболее высокие - в июле (до +17,6˚С). Среднегодовая температура воздуха меняется: в бассейне реки от - 0,3˚С (юг области) до - 2,4˚С (север области).
Территория Томской области характеризуется избыточным увлажнением. Осадки здесь на 100-200мм превышают испарение. Годовая сумма осадков изменяется от 400 до 700мм, 2/3 осадков выпадает в жидком виде в теплое время года (Н.С. Евсеева, А.А. Земцов, 1990).
Летом среднемесячная скорость ветра наименьшая в году и изменяется от 1,6 до 4,3м/с. Зимой скорость ветра колеблется от 2 до 4,8м/с, а осенью - от 1,6 до 4,7м/с. Весна - наиболее ветреный период года, скорость ветра в марте - мае варьирует от 2,4 до 5,1м/с (Ресурсы поверхностных вод СССР, 1973).
По характеру долины и русла Обь условно делят на 3 части:
1. верхняя - от слияния Бии и Катуни до устья реки Томь (986км)
2. средняя - от устья Томи до устья Иртыша (1500км)
3. нижняя - от устья Иртыша до впадения в Обскую губу (1160км).
Скорость течения в среднем варьирует от 0,8 до 2м/с. Длительность половодья составляет 83-140 дней. По характеру уровенного режима Обь относится к Западно-Сибирскому типу (по классификации Б.Д. Зайкова). Для рек этого типа характерно весенне-летнее половодье, сменяющаяся осенне-летней меженью, нарушаемой дождями, и продолжительная зимняя межень. Подъем уровня воды происходит в середине - конце апреля. Ранний подъем бывает в конце марта, а поздний - в начале мая. Средние сроки наступления максимального уровня воды приходится на конец апреля или начало мая. Продолжительность весеннего ледохода колеблется от 1 до 15 суток (А.А. Земцов, 1988).
Воды Оби на участке от устья Томи до устья Иртыша характеризуются по классификации О.А. Алекина как пресные с малой и средней минерализацией (от 70мг/л в период весеннего половодья до 430мг/л в зимнюю межень).
По величине рН воды относятся в среднем к нейтральным, в летне-осенний период - к нейтральным и слабощелочным. Величина рН изменяется от 8,00 до 6,60. Максимальное абсолютное значение рН выявляется в период зимней межени. Это объясняется тем, что кислая и слабокислая реакция водной среды формируется в период открытой воды, когда питание рек осуществляется за счет кислых поверхностно-склоновых и почвенно-грунтовых вод на заболоченных водосборах. В период ледостава единственным источником питания рек становится подземное питание, в результате чего сглаживающее картину поверхностное влияние прекращается, рН возрастает до щелочных значений, характерных для нарушенных участков водотоков преимущественно с рекреационной нагрузкой.
Цветность воды является одним из показателей присутствия растворенного аллохтонного органического вещества. Несмотря на то, что в период половодья основная масса аллохтонной органики поступает с поверхностно-склоновыми водами в водотоки, в этот период наблюдается минимальные значения цветности, что подтверждает разбавляющую роль воды в формировании цветности. В период осенней межени происходит снижение уровня воды с одновременным концентрированием органического вещества. (Е.А. Шорникова, 2007).
Начиная с 50-х гг. ΧΧ века, поверхностные воды Средней Оби испытывают прогрессирующую антропогенную нагрузку, связанную с освоением и эксплуатацией объектов нефтегазового комплекса, интенсивной урбанизацией и созданием обширной и разноплановой инфраструктуры. Характерными загрязняющими веществами бассейна Оби являются соединения железа, меди, цинка, азота аммонийного, нефтепродуктов и фенолов. В наибольших количествах (до 7мг/л и более) в речных водах Оби содержатся Fe, Si и нитрат-анион, в наименьших - фосфат-ион и нитрит-ион. Содержание аммонийного аниона и нитрат-иона являются показателями, по которым качество воды в Оби с вероятностью от 20 до 60% не соответствует установленным нормативам рыбохозяйственного водопользования (О.Г. Савичев, 2003).
Наибольшее содержание железа и марганца наблюдается в зимний и весенний периоды до вскрытия реки, что связано с недостатком кислорода и преобладанием грунтового питания в подледный период (до 30%). Высокое содержание марганца и железа в водах реки обусловлено также и специфическими природными факторами. В подземных водах района характерным является присутствие повышенных концентраций Fe и Mn, что связано с интенсивной заболоченностью территории, насыщенностью органикой и железистыми материалами. Качество воды в средней части реки соответствует ІІ классу качества за счет уменьшения содержания нефтепродуктов, тогда как в нижней части Оби класс качества воды изменяется от "грязной" до "чрезвычайно грязной". В Верхней Оби соответствует III классу качества ("умеренно грязная") (Л.И. Алферова, В.В. Дзюбо, 2005).
Нефтепродукты являются основным видом загрязнения поверхностных вод на территории Среднего Приобья. Минимальная концентрация наблюдается в период половодья. В период осенней и зимней межени, когда расход воды в реке минимален, наблюдается увеличение содержания нефтепродуктов. Присутствие в воде фенолов обусловлено как природными особенностями заболоченных водосборных территорий, так и их поступлением от объектов нефтедобычи в составе нефтесодержащих сточных вод. Максимальное содержание фенолов в воде реки отмечается в период половодья, когда их поступление осуществляется с поверхностно-склоновыми водами (Е.А. Шорникова, 2007).
Помимо антропогенной нагрузки река подвергается действию специфических режимов: длительный ледостав, короткий вегетационный период, низкая концентрация кислорода и т.д. Это могло повлиять на видовой состав фитопланктона и функционирование водоема в целом. К тому же, короткое лето, особенности гидрологического режима (длительный ледостав, растянутое весенне-летнее половодье, преимущественно снеговое питание с заболоченных водосборов) определяет гидрохимические особенности рек таежной зоны Западной Сибири, в том числе и Оби: высокая цветность, пониженные значения рН, высокие концентрации аллохтонного вещества, соединений железа, дефицит растворенного кислорода и т.д.
3. Ценотическая характеристика групп водорослей в 2008 году
При проведении качественной оценки собранного материала было выявлено, что на данном участке реки обитают представители из 4 отделов водорослей (Cyanophyta, Chlorophyta, Bacillariophyta и Euglenophyta),7 классов,14 порядков, 33 семейств,80 родов и 193 видов. Причем первое место по числу видов занимает отдел Chlorophyta (53,8860%). Вторая позиция у Cyanophyta (25,9067%). На третьем и четвертом месте Bacillariophyta и Euglenophyta с 28 и 11 видами, что составляет 14,5078% и 5,95% соответственно (таблица 1).
Таблица 1 - Систематический состав фитопланктона в 2008 г.
| ОТДЕЛ | КЛАСС | ПОРЯДОК | СЕМЕЙСТВО | РОД | ВИД | доля (%) от общего количества видов |
| Chlorophyta | 1 | 4 | 11 | 37 | 104 | 53,8860 |
| Cyanophyta | 3 | 6 | 13 | 22 | 50 | 25,9067 |
| Bacillariophyta | 2 | 3 | 7 | 16 | 28 | 14,5078 |
| Euglenophyta | 1 | 1 | 2 | 5 | 11 | 5,6995 |
| Всего | 7 | 14 | 33 | 80 | 193 | 100 |
Анализируя распределение таксонов водорослей по месяцам, можно отметить, что распределение видов по месяцам неоднородное и колеблется в достаточно больших пределах. Видовое разнообразие представителей разных отделов водорослей на протяжении периода исследований различно. Наибольшее разнообразие диатомовых наблюдалось в июне, августе и сентябре, но представители денного отдела не были найдены в пробах в октябре. Массовое развитие представителей зеленых наблюдалось с июля по сентябрь, сине-зеленых - в июле. Евгленовые были отмечены с июля по сентябрь, но не встречены в мае, июне и октябре. Сине-зеленые не были найден в пробах в июне (таблица 2).
Наибольшее число видов наблюдалось с июля по сентябрь.
Таблица 2 - Распределение видов различных отделов водорослей по месяцам в 2008 г.
| ОТДЕЛ | Диатомовые | Зеленые | Сине-зеленые | Евгленовые | Всего видов |
| МЕСЯЦ | |||||
| Май | 1 | 19 | 3 | 0 | 23 |
| Июнь | 7 | 13 | 0 | 0 | 20 |
| Июль | 4 | 45 | 26 | 2 | 77 |
| Август | 8 | 26 | 14 | 4 | 52 |
| Сентябрь | 9 | 40 | 7 | 2 | 58 |
| Октябрь | 0 | 6 | 2 | 0 | 8 |
Видовой состав фитопланктона у правого берега реки Оби у п. Половинка достаточно разнообразен. Большая часть видов с известной приуроченностью к определенному местообитанию является планктонными обитателями (92 вида из 109, что составляет 84,40%). Представителей донных местообитаний обнаружено всего 10 видов, что составляет 9,18%, перифитона - 7 видов (6,42%). По 43,5% видам не было найдено соответствующей информации.
Среди обнаруженных микроводорослей были найдены виды-индикаторы солености (37 видов, что составляет 19,17%), из которых большую часть составляю виды индифференты (56,76%), такие как Nitzschia gracilis Hantzsch var. minor Skabitsch из диатомовых, Pediastrum simplex Meyen, Actinastrum hantschii Lagerh var. gracile Roll из зеленых и другие. Доля олигогалобов составляет 21,62%, галлофилов - 18,92%. Встречен также 1 вид мезогалоб Synedra tabulata (Ag) Kütz tabulata из диатомовых (2,70%).
По отношению к рН также было обнаружено 32 вида-индикатора (16,58%), из которых 50% составляют индифференты, такие как Pediastrum simplex Meyen, Cosmarium botrytis Menegh var. botrytis, Ulotrix zonata Kűtz. из зеленых и другие.46,88% - ацидофилы (Eremosphaera viridis De Bary, Trachelomonas lacustris var. Sabutata (Skv) Popova, Trachelomonas Matvienkovi Popova из эвгленовых). Обнаружен также 1 вид-алкалифил Nitzschia vermicularis (Kütz) Grun из диатомовых (3,12%) (Приложение В).
Далее был проведен анализ распределения представителей фитопланктона по семействам и выявление родового и видового разнообразия в этих семействах.
Восемь наиболее крупных по числу видов семейств (таблица 3) включают 138 видов из 193, что составляет 71,5025% от общего числа видов. На остальные 25 семейств приходится 55 видов (28,4975%). Таким образом, в видовом составе Оби прослеживается концентрация видов в сравнительно небольшом числе семейств.24 семейства из 33 имеют одно-двухродовой состав и 19 семейств имеют одно-двухвидовое разнообразие. Преобладание маловидовых семейств и родов отличает северную флору и отмечено в составе водорослей Большеземельской тундры, Ямала и Таймыра (В.А. Габышев, 2008).
Наибольшее число семейств характерно для отдела сине-зеленых (13семейств), чуть поменьше - 11 семейств - для отдела зеленых. В отделе диатомовых встречен представители 7 семейств, а в отделе евгленовых - 2 семейства.
Ведущая роль десмидиевых в семейственном спектре Оби отражает голарктические черты флор северного полушария, а также обуславливается мощной подпиткой Оби многочисленными притоками, протекающими по заболоченной территории (Науменко Ю.В., 1996).
К группе показателей систематического разнообразия относятся так называемые пропорции флоры: среднее число видов в семействе (5,85), среднее число родов в семействе (2,42) и среднее число видов в роде (2,41) (В.М. Шмидт, 1984).
Таблица 3 - Крупнейшие по числу видов семейства фитопланктона в 2008 г.
| Место по числу видов | Семейство | Число видов | Число родов | |
| абсолютное | % | |||
| 1 | Desmidiaceae | 36 | 18,6528 | 5 |
| 2 | Oocystaceae | 20 | 10,3627 | 9 |
| 3 | Naviculaceae | 17 | 8,8083 | 7 |
| 4 | Scenedesmaceae | 16 | 8,2902 | 6 |
| 5 | Anabaenaceae | 15 | 7,7720 | 2 |
| 6-7 | Chlorococcaceae | 12 | 6,2176 | 8 |
| 6-7 | Oscilatoriaceae | 12 | 6,2176 | 4 |
| 8 | Euglenaceae | 10 | 5,1813 | 4 |
| Всего | 138 | 71,5025 |
Основу видового богатства отдела Chlorophyta составлял порядок Chlorococcales (53,8462%), включающий семейства Oocystaceae (19,2308%), Scenedesmaceae (15,3846%), Chlorococcaceae (11,5385%), Hydrodictiaceae (5,7692%), Palmellaceae (1,9231%). На втором месте по видовому разнообразию порядок Zygnematales (39,4230%), содержащий в своем составе представителей семейств Desmidiaceae (34,6153%), Mesotaeniaceae (2,8846%), Zygnemataceae (1,9231%). Среди 11 семейств 5 имеют двувидовой состав, что составляет 7,69% видового разнообразия (таблица 4).
Таблица 4 - Семейства отдела Chlorophyta альгофлоры
Средней Оби в 2008 г.
| СЕМЕЙСТВО | ЧИСЛО ВИДОВ | ЧИСЛО РОДОВ | Процентное отношение видового богатства семейств (в%) |
| 1. Chlorococcaceae | 12 | 8 | 11,5385 |
| 2. Chlamydomonadaceae | 2 | 1 | 1,9231 |
| 3. Desmidiaceae | 36 | 5 | 34,6153 |
| 4. Hydrodictiaceae | 6 | 1 | 5,7692 |
| 5. Mesotaeniaceae | 3 | 2 | 2,8846 |
| 6. Oocystaceae | 20 | 9 | 19,2308 |
| 7. Palmellaceae | 2 | 1 | 1,9231 |
| 8. Scenedesmaceae | 16 | 6 | 15,3846 |
| 9. Ulothricaceae | 3 | 1 | 2,8846 |
| 10. Volvocaceae | 2 | 1 | 1,9231 |
| 11. Zygnemataceae | 2 | 2 | 1,9231 |
| Всего | 104 | 37 | 100 |
Почти все видовое разнообразие отдела Bacillariophyta обеспечивает класс Pennatophyceae (96,4286%), Centrophyceae составляют оставшиеся 3,5714%. В классе Pennatophyceae наиболее широко представлены виды порядка Raphales (78,5715%), включающего в себя семейства Naviculaceae (60,7143%), Epithemiaceae (7,1430%), Achnanthaceae (3,5714%), Eunotiaceae (3,5714%), Nitzschiaceae (3,5714%).
Среди 7 встреченных семейств диатомовых 5 имеют одно-двухвидовой состав, что составляет 21,43% от видового богатства отдела Bacillariophyta. Оставшиеся 2 семейства (Naviculaceae и Fragillariaceae) составляют 78,57% (таблица 5).
Таблица 5 - Семейства отдела Bacillariophyta альгофлоры
Средней Оби в 2008 г.
| СЕМЕЙСТВО | ЧИСЛО ВИДОВ | ЧИСЛО РОДОВ | Процентное отношение видового богатства семейств (в%) |
| 1. Achnanthaceae | 1 | 1 | 3,5714 |
| 2. Coscinodiscaceae | 1 | 1 | 3,5714 |
| 3. Eunotiaceae | 1 | 1 | 3,5714 |
| 4. Epithemiaceae | 2 | 2 | 7,1430 |
| 5. Fragillariaceae | 5 | 4 | 17,8571 |
| 6. Naviculaceae | 17 | 7 | 60,7143 |
| 7. Nitzschiaceae | 1 | 1 | 3,5714 |
| Всего | 28 | 17 | 100 |
Основу видового богатства отдела Cyanophyta обеспечивают 2 класса Chroococcophyceae (32%) и Hormogoniphyceae (62%). В классе Chroococcophyceae все представители принадлежат порядоку Chroococcales. В классе Hormogoniphyceae наиболее широко представлен порядок Nostocales (34%), включающий семейства Anabaenaceae (30%), Oscilatoriaceae (24%), Schizotrichaceae (2%), Nostocaceae (2%), Rivulariaceae (2%).
В структуре отдела Cyanophyta представлено много одно - и двухвидовых семейств, видовой состав которых составляет 24% от обнаруженных на данном участке Оби. Таким образом, остальные 4 семейства из 13 содержат в себе 76% видового разнообразия растительного планктона.
Таблица 6 - Семейства отдела Cyanophyta в альгофлоре
Средней Оби в 2008 г.
| СЕМЕЙСТВО | ЧИСЛО ВИДОВ | ЧИСЛО РОДОВ | Процентное отношение видового богатства семейств (в%) |
| 1. Anabaenaceae | 15 | 2 | 30 |
| 2. Chamaesiphonaceae | 1 | 1 | 2 |
| 3. Coccobactreaceae | 6 | 4 | 12 |
| 4. Coelosphaeriaceae | 1 | 1 | 2 |
| 5. Gloeocapsaceae | 2 | 1 | 4 |
| 6. Merismopediaceae | 2 | 1 | 4 |
| 7. Microcystidaceae | 5 | 2 | 10 |
| 8. Nostocaceae | 1 | 1 | 2 |
| 9. Oscilatoriaceae | 12 | 4 | 24 |
| 10. Pleurocapsaceae | 2 | 1 | 4 |
| 11. Rivulariaceae | 1 | 1 | 2 |
| 12. Schizotrichaceae | 1 | 1 | 2 |
| 13. Stigonemataceae | 1 | 1 | 2 |
| Всего | 50 | 21 | 10 |
Все представители отдела Euglenophyta принадлежат одному порядку Euglenales и двум семействам в его составе Anabaenaceae (90,909%) и Astasiaceae (9,091%) (таблица 7).
Таблица 7 - Семейства отдела Euglenophyta в альгофлоре Средней Оби в 2008 г.
| СЕМЕЙСТВО | ЧИСЛО ВИДОВ | ЧИСЛО РОДОВ | Процентное отношение видового богатства семейств (в%) |
| 1. Astasiaceae | 1 | 1 | 9,091 |
| 2. Anabaenaceae | 10 | 4 | 90,9090 |
| Всего | 11 | 5 | 100 |
Расположение ведущих по числу видов родов на данном участке Оби у п. Половинка свидетельствует о том, что 8 родов из 80 включают 41,9689%, от общего числа видов (таблица 8). Таким образом, оставшиеся 72 вида составляют 58,0311%. Двенадцать родов в исследуемом сообществе является двуродовыми (15%), 43 рода одновидовыми (53,75%).
Если рассматривать распределение видов по родам, то также видна концентрация видов в небольшом их количестве (в 9 родах из 80-ти содержится 41,9689% видов).
Таблица 8 - Крупнейшие по числу видов роды фитопланктона в 2008 г.
| Место по числу видов | Род | Число видов | % |
| 1 | Closterium | 20 | 10,3627 |
| 2 | Anabaena | 14 | 7,2539 |
| 3 | Scenedesmus | 9 | 4,6632 |
| 4 | Ankistrodesmus | 8 | 4,1451 |
| 5 | Oscillatoria | 7 | 3,6269 |
| 6-8 | Pediastrum | 6 | 3,1088 |
| 6-8 | Pleurotaenium | 6 | 3,1088 |
| 6-8 | Navicula | 6 | 3,1088 |
| 9 | Trachelomonas | 5 | 2,5907 |
| Всего | 81 | 41,9689 |
Фитопланктон влияет на скорость круговорота веществ в водоеме. В зависимости от своих физиологических особенностей водоросли накапливают те или иные химические элементы с большей или меньшей интенсивностью. Изъятие химических элементов из водной среды может происходить в разный срок, который в значительной степени определяется морфологией водорослей: виды, обладающие мягкой слизистой оболочкой, после отмирания минерализуются непосредственно в толще воды, что обеспечивает быстрый круговорот веществ. Панцирные формы оседают на дно и там медленно и не полностью минерализуются, т.е. возврат химических элементов в воде замедляется, а часть из них выпадает из круговорота веществ (Г.М. Лаврентьева, 1984г).
При анализе фитопланктона следует уделять внимание редким видам, так как они являются индикаторными для условий данного водоема (Приложение С).
4. Характеристика биотопов Средней Оби по микроводорослям в 2008г
Как известно, на видовое разнообразие фитопланктона и его количественное развитие влияют многие факторы среды обитания, такие как: рН воды, ее температура, наличие в воде растворенных веществ, содержание кислорода и т.д.
Материалом для выяснения влияния условий местообитания на развитие водорослей послужили 5 проб, собранных 27.07.08г в течение 80 минут (для меньшего влияния изменяющихся погодных условий со временем) на различных участках реки Обь, расположенных на небольшом отдалении друг от друга.
При сравнении видового состава фитопланктона в данных биотопах было обнаружено 65видов, из которых 11 общих для 5 отобранных проб. Все эти виды относились к отделу Chlorophyta. Четыре вида из них являются индикаторами сапробности Crucigenia tetrapedia (Kirhn) W. et. G West, Actinastrum hantschii Lagerh var. gracile Roll, Tetraëdron minutum (A. Br) Hansg f. minutum и Scenedesmus acuminatus (Legerh) Chod. Первые три являются β-мезосапробами, а последний - 0-β-мезосапроб.
Менее всего сходства в видовом разнообразии в пробах с 3-го участка в силу малого числа обнаруженных видов. В пробах на этом участке присутствует 2 вида из отдела Chlorophyta, 2 вида из сине-зеленых и 1 вид из евгленовых. Данный участок характеризуется отсутствием течения, илисто-песчаным дном, небольшой глубиной (до 30см). По берегу и на дне произрастают высшая водная растительность, вода прогрелась до 30˚С.
В первой пробе, взятой с участка, характеризующегося глинисто-песчаным дном, наличием течения, температурой воды 24,4˚С, прозрачностью в 40см, обнаружены представители отдела Chlorophyta (17 видов), Bacillariophyta (1 вид) и отдела Cyanophyta (3 вида).
Во второй пробе, взятой с небольшой отмели на середине реки, где практически отсутствует течение, дно песчано-глинистое, нет растительности, с температурой воды 25˚С, прозрачностью в 60см, обнаружен 1 вид из евгленовых, 2 вида из отдела Cyanophyta и 13 видов зеленых водорослей.
Четвертая проба взята с правого берега реки, на котором произрастают древесные породы и осоковые. Проба бралась у берега с каменистым дном. Течение на данном участке слабое, температура воды 25˚С, прозрачность - 45см. В ней обнаружены представители отделов Bacillariophyta (1 вид), Cyanophyta (4 вида) и Chlorophyta (14 видов).
Пятая проба взята в месте разделения реки островом на два рукава, на берегу которого произрастают в основном кустарниковые и ивы, осоковые и злаки. Проба отбиралась на участке с песчано-глинистым дном, с температурой воды 22,4˚С и прозрачностью в 50см. В пробах обнаружены были виды из отдела Euglenophyta (3 вида), Bacillariophyta (1 вид), Cyanophyta (3 вида) и Chlorophyta (14 видов).
Вне зависимости от условий обитания число видов в пробах сравнимо между собой, за исключением пробы, взятой с 3-го участка, где наблюдается их минимальное число. Наибольшее развитие фитопланктона наблюдается на 1 и 2 участках (2,076-2,33*109кл/м³), немного меньше на 4 и5 (1,512-1,575*109кл/м³) и минимальный уровень вегетации обнаружен на 3-м участке (0,315*109кл/м³).
При анализе видов индикаторов сапробности было выявлено, что все участки относятся к β-мезосапробной зоне или переходной от α-олигосапробной к β-мезосапробной зоне.
Таким образом, анализ проб с разных участков реки показал, что видовое разнообразие фитопланктона довольно однородно, а скорость вегетации отличается в зависимости от условий данного участка. Наиболее благоприятными в этом отношении оказались 1 и 2 участок.
Диаграмма 1 - Зависимость между численностью фитопланктона и условиями местообитания
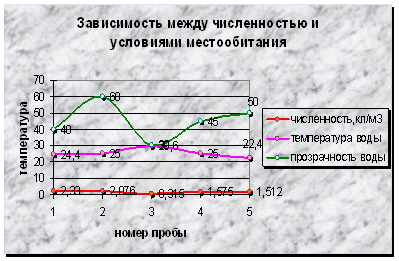
Данная диаграмма показывает отсутствие зависимости численности фитопланктона от температуры участка реки и прозрачности воды. Скорее всего, уровень вегетации зависит от скорости течения, грунта, высшей водной растительности, гидрохимии.
5. Сезонное развитие фитопланктона Средней Оби в 2008 г.
При анализе проб фитопланктона за 6 месяцев, просматривается доминирование по численности отдела Chlorophyta, на втором и третьем месте соответственно стоят отделы Cyanophyta и Bacillariophyta. За ними следует Euglenophyta. (таблица 9).
Таблица 9 - Сезонное развитие микроводорослей разных
отделов фитопланктона в 2008г.
|
Численность отдела, *109экз. /м3 |
Chlorophyta | Cyanophyta | Bacillariophyta | Euglenophyta | Всего | Средняя численность |
| месяц | ||||||
| МАЙ |
0,630 |
0,126 | 0,063 | 0 | 0,819 | 1,008 |
|
1,134 |
0,063 | 0 | 0 | 1, 197 | ||
| ИЮНЬ |
0,692 |
0 | 0,315 | 0 | 1,007 | 1,007 |
|
0,692 |
0 | 0,315 | 0 | 1,007 | ||
| ИЮЛЬ |
0,943 |
0,189 | 0 | 0,063 | 1, 195 | 1,4849 |
|
1,511 |
1, 196 | 0,063 | 0 | 2,770 | ||
| 0,8493 |
0,6604 |
0,0943 | 0 | 1,604 | ||
|
0,7234 |
0, 2044 | 0,1101 | 0,0629 | 1,1008 | ||
|
0,5191 |
0,236 | 0 | 0 | 0,7551 | ||
| АВГУСТ |
0,3775 |
0,3302 | 0 | 0 | 0,7077 | 1,1691 |
|
0,3776 |
0 | 0,2831 | 0,0944 | 0,7551 | ||
| 0,2359 |
0,6134 |
0,0944 | 0 | 0,9437 | ||
|
1,262 |
0,630 | 0,252 |
0,126 |
2,270 | ||
| СЕНТЯБРЬ |
0,813 |
0,063 | 0,252 | 0,252 | 1,380 | 2,525 |
| 0,57 |
0,88 |
0 | 0 | 1,45 | ||
|
3,69 |
0,5077 | 0,5483 | 0 | 4,7456 | ||
| ОКТЯБРЬ | 0,1888 |
0,3301 |
0 | 0 | 0,5189 | 0,3774 |
| 0,0944 |
0,1415 |
0 | 0 | 0,2359 |
Из таблицы видно, что наименьший уровень эвтрофикации в водоеме наблюдался в октябре, а максимума достиг в сентябре при массовом развитии зеленых.
В течение почти всего периода вегетации прослеживается сильная вегетация представителей отдела зеленых. Доминирование зеленых наблюдается с мая по сентябрь, только в октябре численность сине-зеленых немного превышала развитие представителей отдела Chlorophyta.
Данные по численности фитопланктона, полученные при анализе распределения ее по месяцам, можно представить в виде диаграммы (диаграмма 2).
Диаграмма 2 - сезонная динамика фитопланктона в 2008 году
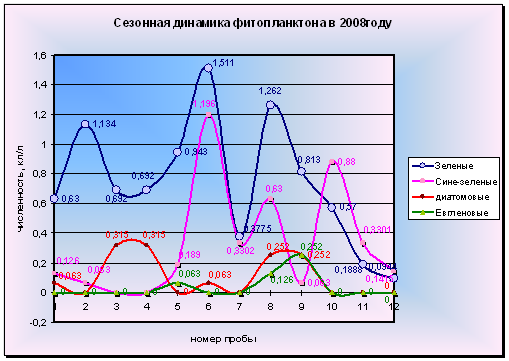
В общем можно заметить одновременные периодические всплески численности сразу нескольких отделов микроводорослей. Явный пик вегетации зеленых водорослей хорошо прослеживается на диаграмме. Немного смещены от него во времени небольшие всплески численности сине-зеленых и диатомовых. Также наблюдается небольшой подъем численности зеленых и сине-зеленых в июне с опережающей их вспышкой численности диатомовых.
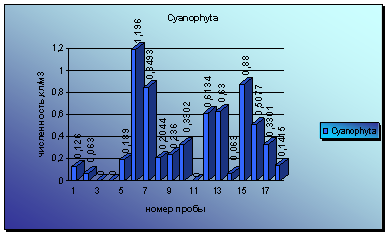
При рассмотрении динамики численности представителей отдела сине-зеленых, можно отметить 2 крупные вспышки численности. Первая зарегистрирована при взятии пробы 18 июля, но имела кратковременный характер и быстро угасла уже к концу июля. Она была вызвана развитием водорослей родов Anabaena и Phormidium.
Вторая вспышка обнаружена в пробах за 14 сентября и вызвана вегетацией только водорослей р. Anabaena. Численность сине-зеленых в этот раз превышала численность зеленых, но также недолго. Вскоре зеленые достигли своего максимума вегетации, а сине-зеленые начали медленными темпами снижать свои темпы размножения. Но еще к концу октября наблюдается довольно сильная вегетация представителей этого отдела.
В видовом отношении не происходит постепенной замены одних видов другими, и можно обнаружить, среди водорослей майской альгофлоры виды, преобладающие в октябре (диаграмма 3).
Диаграмма 3 - Сезонная динамика Cyanophytа в 2008 г.
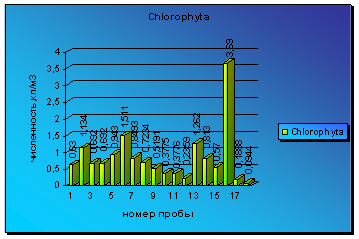
Анализируя колебания численности зеленых микроводорослей, можно отметить 3 небольших и один крупный всплеск вегетации водорослей. Первый подъем наблюдается уже в середине мая, затем уровень вегетации немного снижается до следующего пика численности в середине июля. Первый подъем был вызван водорослями родов Closterium и Pleurotaenium, второй - развитием водорослей родов Closterium и Palmella. Затем наблюдается медленный и постепенный спад вегетации, который сходит почти до нуля. Но в середине августа наблюдается резкий скачок численности, который постепенно к концу сентября переходит в еще один всплеск численности, самый крупный за весь период наблюдений вегетации всех отделов водорослей. Он был вызван в основном развитием водорослей р. Ankistrodesmus и Closterium (диаграмма 4).
Диаграмма 4 - Сезонная динамика Chlorophyta в 2008 г.
Колебания численности диатомовых проявляются хорошо и периодически снижаются до нуля. Из 18 проанализированных проб в 7 не было обнаружено водорослей данного отдела. Достаточно высокий уровень вегетации диатомовых был зарегистрирован в июньских пробах. В этот период наблюдалась вегетация видов Asterionella formosa Hass, Fragillaria construens (Ehr) Grum var. Subsalina Hust, Pinnularia bigibba Ehr. Var. mesogondyla f. interrupta, Synedra tabulata (Ag) Kütz tabulate и Melosira islandica O. Müll subsp. islandica Hust. Далее, с июня по август, встречались лишь отдельные представители отдела диатомовых, не вносящих большого вклада в общий уровень вегетации водорослей. Только в начале сентября вновь повысился уровень вегетации диатомовых. Он продержался на невысоком уровне до конца сентября и вылился в самую крупную вспышку численности диатомовых, вызванную развитием представителей родов Amphora, Opephora, Pinnularia и Navicula (диаграмма 5).
Таким образом, вегетация диатомовых за весь период наблюдений не достигала высоких уровней, а периодами условия были неблагоприятными для развития представителей отдела Bacillariophyta, что вызывало полное их отсутствие в исследуемых пробах.
Часто лимитирующим фактором для диатомовых является содержание в воде Si. По мнению большинства исследователей, для того, чтобы наступило "цветение" водоема за счет бурного развития диатомовых, необходимо содержание кремния в воде не менее 4мг/л. Однако в природе часто и при меньших концентрациях Si диатомовые водоросли продолжают развиваться. По отношению к кислотности среды диатомовые более приурочены к кислым водам (О.Г. Воропаева, 1988г). Но, по-видимому, температурный режим, химический состав воды и другие фактор помешали нормальному развитию водорослей.
Диаграмма 5 - Сезонная динамика Bacillariophyta в 2008 г.
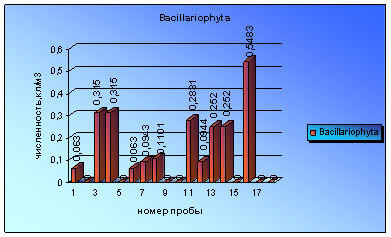
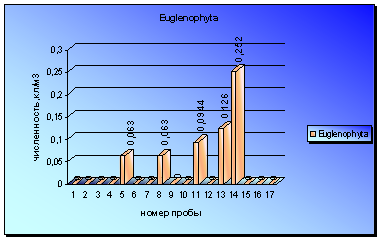
Представители отдела Euglenophyta встречены в пробах еще реже диатомовых. Только в 5 пробах были зарегистрированы небольшие вспышки численности евгленовых. Впервые микроводоросли данного отдела были обнаружены в пробе за 3 июля и периодически встречались до начала сентября (диаграмма 6).
На протяжении этого периода можно отметить незначительные увеличения численности, которые достигли наибольшей величины в пробах за 1 сентября. Максимальный всплеск численности водорослей был вызван интенсивной вегетацией видов Trachelomonas lacustris Drez. Emend Balech и Trachelomonas lacustris var. Sabutata (Skv) Popova.
Видовой состав евгленовых на протяжении всего времени их вегетации постоянно менялся. И невозможно найти одни и те же виды, встречающиеся в разные периоды времени.
Диаграмма 6 - Сезонная динамика Euglenophyta в 2008 г.
6. Определение сапробности исследуемого участка реки Обь в 2008 году
В ходе исследований в 2008 году нами было выявлено 29 видов-индикаторов сапробности. Из них 16 видов зеленых, 7 видов диатомовых, 5 - эвгленовые и 2 вида сине-зеленые. Чаще всего встречались виды, характерные для β-мезосапробной (13 видов) и олигосапробной (9 видов) зон. Обнаружено также 4 вида, характерных для 0-β-сапрбной зоны, 1 вид - для β-α-мезосапробной зоны и 3 вида α-мезосапробной зоны (таблица 10).
Таблица 10 - Виды - индикаторы сапробности.
| отдел | вид | сапробность | |
| Chlorophyta | Ulotrix zonata Kűtz | 0 | |
| Bacillariophyta | Amphora ovalis Kütz var. pediculus Kütz | 0 | |
| Bacillariophyta | Asterionella formosa Hass | 0 | |
| Bacillariophyta | Navicula tuscula (Ehr) Grun | 0 | |
| Bacillariophyta | Fragillaria construens (Ehr) Grum var. Subsalina Hust | 0 | |
| Bacillariophyta | Synedra tabulata (Ag) Kütz tabulata | 0 | |
| Bacillariophyta | Nitzschia vermicularis (Kűtz) Grun | 0 | |
| Cyanophyta | Gloeocapsa minuta (Kűtz) Hollerb ampl | 0 | |
| Bacillariophyta | Nitzschia gracilis Hant | 0 | |
| Cyanophyta | Anabaena spiroides Kleb | 0-β | |
| Chlorophyta | Scenedesmus acuminatus (Legerh) Chod | 0-β | |
| Euglenophyta | Euglena granulata (Klebs) Schmitz | 0-β | |
| Euglenophyta | Lepocinclis fusiformes (Carter) Lemm | 0-β | |
| Chlorophyta | Crucigenia tetrapedia (Kirhn) W. et. G West | β | |
| Chlorophyta | Gonium sociale Warm | β | |
| Cyanophyta | Microcystis pulverea (Wood) Forti emend f. parasitica (Kűtz) Elenk | β | |
| Chlorophyta | Closterium moniliferum (Bory) Ehr. | β | |
| Chlorophyta | Pandorina morum (Müll) Bory | β | |
| Chlorophyta | Pediastrum simplex Meyen | β | |
| Chlorophyta | Pediastrum dulex Meyen | β | |
| Chlorophyta | Pediastrum tetras var. tetras | β | |
| Chlorophyta | Tetraëdron caudatum (Сorda) Hansg | β | |
| Chlorophyta | Tetraëdron minutum (A. Br) Hansg | β | |
| Chlorophyta | Actinastrum hantschii Lagerh | β | |
| Chlorophyta | Anristrodesmus acicularis Korschik | β | |
| Chlorophyta | Closterium parvulum Näg | β | |
| Cyanophyta | Merismopedia tenuissima Lemm | β-α | |
| Cyanophyta | Oscillatoria splendida Grev | α | |
| Chlorophyta | Closterium acerosum (Schr) Ehr. | α | |
| Chlorophyta | Cosmarium botrytis Menegh | α | |
Анализ обнаруженных видов-индикаторов и расчет сапробности микроводорослей позволил выявить следующее (таблица 11).
Таблица 11 - Сапробность участка правого берега Оби по сезонам в 2008 г.
| Месяц | Индекс Пантле и Букк | сапробность |
| Май | 2,50 | α-β-мезосапробная |
| Июнь | 1,40 | олигосапробная |
| Июль | 2,074 | β-мезосапробная |
| Август | 1,903 | β-мезосапробная |
| Сентябрь | 2, 197 | β-мезосапробная |
| Октябрь | 2,290 | β-мезосапробная |
| Среднее значение | 2,061 | β-мезосапробная |
Лишь июньский фитопланктон характеризует исследуемый участок реки как олигосапробный. С июля по октябрь сапробность воды повышалась до β-мезосапробной, а максимум приходится на май с массовым развитием зеленых водорослей. Изменение величин сапробности незначительны, что вероятно можно рассматривать как признак довольно высокой стабильности экосистемы реки, пока самостоятельно справляющейся с возрастающим загрязнением (диаграмма 7). Возможно, низкий уровень сапробности обусловлен и погодными условиями в данный период. Для этих сезонов был характерен повышенный уровень воды, что могло повлиять на снижение концентрации фитопланктона в воде. Майское загрязнение может быть связано со сбросом вод из водохранилища в поводок.
Диаграмма 7 - Сезонное изменение средних величин сапробности Средней Оби у п. Половинка
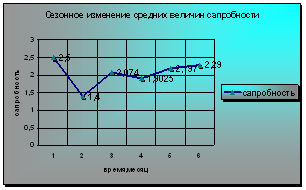
Полученные результаты позволяют отнести обследуемый участок Средней Оби к водам удовлетворительной чистоты (β-мезосапробной зоне).
7. Сравнение динамики развития фитопланктона за 2007-2008 года
В качестве сравниваемого материала служат 7 проб, взятых в 2007 году и 18 проб 2008 года, собранных в период с мая по октябрь.
В мае 2007 года наблюдалось более интенсивное развитие фитопланктона. По сравнению с численностью водорослей 1,95*109кл/м³ в 2007г в начале и середине месяца в 2008 году наблюдался уровень вегетации 0,819*109 и 1, 197*109кл/м³ соответственно. Также различие проявляется и в доминировании водорослей: в 2007г по численности преобладал отдел Bacillariophyta, в 2008г - отдел Chlorophyta. Общих видов в сравниваемые периоды не обнаружено, но встречаются рода Pleurotaenium и Anristrodesmus из отдела зеленых водорослей. В прошлом году в мае вегетировали зеленые, диатомовые и сине-зеленые водоросли, так же, как и в этом году, но в разных пропорциях.
Июнь 2007 года так же отличается более интенсивным развитием микроводорослей по сравнению с 2008 годом. Численность фитопланктона в июне пошлого года составляет 1,64*109кл/м³, в 2008г - 1,007*109кл/м³. Июнь 2007г отличался достаточно однородным развитием представителей отделов Bacillariophyta, Chlorophyta, Cyanophyta и Euglenophyta с небольшим преобладание отдела Euglenophyta. В 2008 году в водоеме были представлены виды отделов Chlorophyta и Bacillariophyta с доминированием представителей отдела Chlorophyta. Общим видом в данный период был Actinastrum hantschii Lagerh var. gracile Roll, также встречались представители родов Crucigenia и Scenedesmus из отдела зеленых водорослей.
Середина июля 2008 года по интенсивности развития водорослей сравнима с этим же периодом в 2007г (2,77*109кл/м³). Тогда как в начале и конце месяца наблюдается ослабление развития водорослей с плотностью 1, 195 и 0,7551*109кл/м³. В течение обоих периодов наблюдалось развитие водорослей из отделов Cyanophyta, Chlorophyta и Bacillariophyta, но в прошлом году доминировал отдел Cyanophyta, а в 2008г - представители отдела Chlorophyta. В июле 2007 и 2008гг в пробах встречены представители родов Anabaena (Cyanophyta), Crucigenia (Chlorophyta) и вид Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta).
В августе 2007 года и в первых двух декадах августа 2008г наблюдается незначительное развитие водорослей, в пределах 0,7-0,94*109кл/м³, а к концу месяца в 2008г отмечено увеличение вегетации микроводорослей. В течение обоих периодов интенсивно размножались водоросли из отделов Cyanophyta, Chlorophyta. Но в 2007г встречено несколько представителей отдела Euglenophyta, а в 2008г - отдела Bacillariophyta. В пробах за данный период были отмечены общие виды Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta), Crucigenia tetrapedia (Kirhn) W. et. G West (Chlorophyta) и Scenedesmus acuminatus (Legerh) Chod (Chlorophyta).
В сентябре 2007г и начале месяца 2008г плотность водорослей не имела больших различий и находилась в пределах 1,32 - 1,45*109кл/м³. В 2008 году к середине сентября было отмечено резкое повышение развития водорослей до 4,7456*109кл/м³ в основном за счет большого количества водорослей из отдела Chlorophyta. На протяжении обоих периодов наблюдалась вегетация водорослей из отделов Bacillariophyta, Chlorophyta, Cyanophyta и Euglenophyta. В 2007 году доминировал отдел Chlorophyta, а в сентябре 2008 года наблюдалась первоначально усиленная вегетация представителей из отделов Euglenophyta и Chlorophyta, затем их место заняли водоросли из отдела Cyanophyta, а к концу месяца вновь преобладали представители из отдела зеленых водорослей. В пробах, за эти периоды исследований, встречались виды Scenedesmus bijugatus Kűtz, Actinastrum hantschii Lagerh var. gracile Roll, а также представители родов Crucigenia, Pediastrum (Chlorophyta), Anabaena (Cyanophyta) и Trachelomonas (Euglenophyta).
Октябрь 2008 года отличается меньшей вегетацией фитопланктона (0,5189 - 0,2359*109кл/м³) по сравнению с аналогичным периодом в прошлом году (1,32*109кл/м³). Альгофлора октября 2007г более разнообразно представлена: встречены представители отделов Bacillariophyta, Chlorophyta, Cyanophyta и Euglenophyta и Chrysophyta. Тогда как в октябре 2008г обнаружены виды отделов Chlorophyta и Cyanophyta. Общим является только один вид из рода Closterium.
Рассматривая общую картину динамики фитопланктона в 2007-2008гг, можно отметить, что в среднем уровень вегетации микроводорослей в 2008 году превышает численность фитопланктона в 2007 году лишь в августе и сентябре.
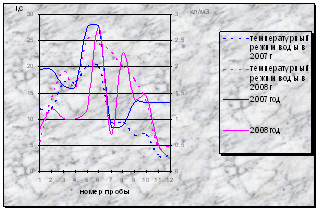
Анализ температурного режима воды, показал, что в 2008 году он заметно превышал уровень температур в то же время 2007 года, кроме октября 2007 года, когда температура воды была немного выше, чем в аналогичный период в 2008 году.
В октябре 2007 года основной вклад в вегетирующий состав микроводорослей внесли холодолюбивые виды отдела Bacillariophyta и теплолюбивые водоросли отдела Cyanophyta. В октябре 2008 года наибольшего развития достигли водоросли из отдела Cyanophyta, хотя температура вод не превышала 5˚С, что более благоприятно для развития холодолюбивых видов, таких как диатомовые.
В августе и сентябре 2008 года развитие фитопланктона значительно превышает уровень вегетации водорослей за этот период 2007 года. Также разрыв температурного диапазона между 2008 и 2007 годом - максимальный. Температура воды в 2008 году намного превышает таковую в 2007 году, что, возможно, и обусловило высокий уровень вегетации водорослей в данном периоде в 2008 году (диаграмма 8).
Диаграмма 8 - Сравнение динамики численности фитопланктона и температурного режима в 2007-2008гг
Численность зеленых водорослей в мае, июне и сентябре 2008 года превышает уровень вегетации представителей этого отдела за тот же период 2007 года. Но максимумы их развития практически совпадают во времени и приурочены к сентябрю. Можно отметить, что всплеск численности в 2008 году был более сильным, чем в 2007 году, что обусловлено более благоприятным температурным режимом в 2008 году.
Минимальный уровень вегетации наблюдался в октябре обоих годов, что связано с наиболее низкими температурами в этот период (диаграмма 9).
Диаграмма 9 - Динамика численности зеленых водорослей в 2007-2008гг
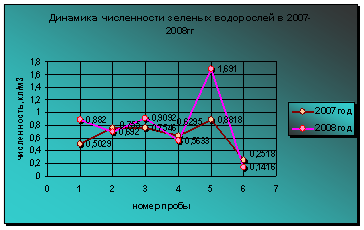
Первую половину периода исследований фитопланктона, до августа 2007 года, доминировал по численности комплекс сине-зеленых. С августа по октябрь 2008 года наибольший уровень развития показал этот же комплекс сине-зеленых. Максимумы развития сине-зеленых приурочены в 2007 и 2008 году к июлю. Наибольшее развитие сине-зеленых отмечено в 2007 году, хотя температура воды в этот период в 2007 году была ниже, чем в 2008 году (диаграмма 10).
Минимальные уровни развития представителей сине-зеленых приурочены к разным периодам. В 2007 году наиболее низкий уровень вегетации водорослей данного отдела отмечен в августе, а в 2008 году - в июне. В августе 2007 года из сине-зеленых был встречен лишь один вид Anabaena Hassalii (Kűtz) Wittr, который может развиваться при широком диапазоне температур.
Диаграмма 10 - Динамика численности сине-зеленых водорослей в 2007-2008гг
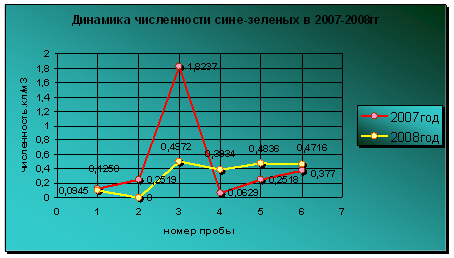
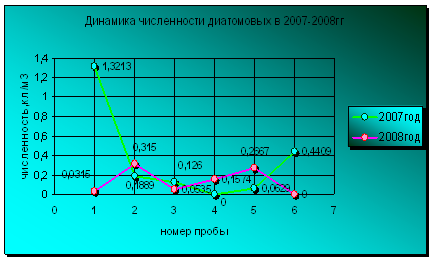
Динамика диатомовых водорослей в сравниваемые годы совершенно различается. Уровень вегетации представителей отдела диатомовых был выше в июне, августе и сентябре 2008 года по сравнении с 2007 годом. Колебания численности диатомовых в 2007 году имеет большую амплитуду по сравнению с уровнем развития этих водорослей в 2008 году, который не имел особых максимумов развития в течение всего периода вегетации (диаграмма 11). В 2007 году пик развития диатомовых водорослей приходится на май, а в 2008 году на июнь. Но уровень вегетации диатомовых в прошлом году намного превышает предельный уровень численности представителей данного отдела в 2008 году, хотя температурный режим мая-июня двух лет не имеет больших отличий. Полностью отсутствовали представители диатомовых в августе 2007 года и октябре 2008 года. Эти периоды характеризовались низкими температурами.
Диаграмма 11 - Динамика численности диатомовых в 2007-2008 гг.
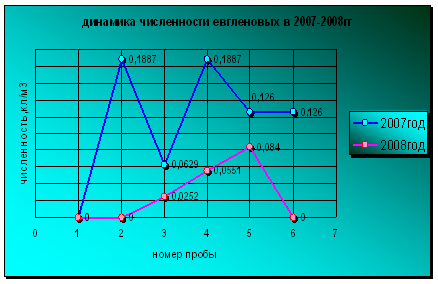
Развитие евгленовых в 2007 году значительно превышает уровень вегетации водорослей в 2008 году и имеет 2 пика численности в июне и августе. В 2008 году евгленовые встречались всего на протяжении трех месяцев: с июля по сентябрь с постепенным возрастанием численности и достижения своего пика развития в сентябре (диаграмма 12).
Диаграмма 12 Динамика численности евгленовых в 2007-2008гг
Таким образом, можно сказать, что в целом динамика численности микроводорослей в 2007-2008 годах имела сходный характер. Вспышки и спады вегетации фитопланктона приурочены к примерно одним и тем же периодам. Но при рассмотрении динамики развития каждого отдела водорослей в отдельности видны явные различия во времени пиков и спадов их вегетации.
Заключение
1. - На обследуемом участке реки Обь нами выявлено 193 вида, принадлежащих к 4 отделам, 7 классам, 14 порядкам, 33 семействам и 80 родам.
2 - Анализ экологии микроводорослей показал, что преобладают планктонные обитатели (84,40%), индифференты по солености (56,76%), индифференты по рН (50%).
3 - На основе определения индикаторных видов была определена сапробность исследуемого участка реки по методу Пантле и Букк. Выявлено, что лишь в июне вода в реке характеризовалась как олигосапробная, на протяжении остального времени - β-мезосапробная.
4 - Распределение микроводорослей по месяцам в 2007 году носило более однородный характер, тогда как в 2008 году количество встречаемых видов неоднородно (максимально с июля по сентябрь).
5 - Водоросли всех отделов имели максимумы развития в разное время и вносили неравный вклад в альгоценоз исследуемого участка Средней оби, но в целом, больший вклад в вегетирующий комплекс фитопланктона внесли представители отдела Chlorophyta.
Список используемой литературы
1. Алферова Л.И. Томская область: водные ресурсы, водохозяйственная деятельность и проблемы питьевого водоснабжения населения / Л.И. Алферова, Дзюбо В.В. // Водное хозяйство России. - 2005. - т.7, №4. - С.333-345
2. Габышев В.А. Водоросли планктона водоемов бассейна реки Молодо (Россия, Якутия) // Гидробиологический журнал. - 2008. - т.44. - №3. - С.12-18.
3. География Томской области / А.А. Земцов [и др.]. - Томск: изд-во Том. ун-та, 1988. - 246с.
4. Догановский А.М. Гидросфера земли / А.М. Догановский, Н.В. Малинин - С. - П.: изд-во Гидрометеоиздат, 2004. - 630с.
5. Евсеева Н.С., Земцов А.А. Рельефообразование в лесоболотной зоне западно-Сибирской равнины / Н.С. Евсеева, А.А. Земцов Томск: изд-во ТГУ, 1990. - 240с.
6.Л.Е. Комаренко Пресноводные диатомовые и сине-зеленые водоросли водоемов Якутии / Л.Е. Комаренко, И.И. Васильева. - М.: изд-во Наука, 1975. - 423с.
7.Л.Е. Комаренко Пресноводные зеленые водоросли водоемов Якутии / Комаренко Л.Е., Васильева И.И. -М.: изд-во Наука, 1978. - 284с.
8.В.И. Кузьмичева Погрешность метода склянок при определении продукции и деструкции планктона // биология внутренних вод. - 1970. - №7. - С.17-20.
9. Малик Л.К. Гидрологические проблемы преобразования природы Западной Сибири / Л.К. Малик. - Л.: Гидрометеоиздат, 1978. - 180с.
10. Матвиенко А.М. Определитель пресноводных водорослей СССР. Золотистые водоросли / Матвиенко А.М. -М.: изд-во Советская наука, 1954. - Вып.3. - 188с.
11. Методические рекомендации по сбору и обработке при гидробиологических исследованиях на пресноводных водоемах. Фитопланктон и его продукция / Лаврентьева Г.М., Винберг Г.Г. - Л.: ГосНИОРХ, 1984. - 32с.
12. Методы физиолого-биохимического исследования водорослей в гидробиологической практике/ Л.А. Сиренко [и др.]. - Киев: изд-во Наукова думка, 1975. - 247с.
13. Науменко Ю.В. Фитопланктон реки Оби: автореферат на соискание ученой степени ДБН.03.00-05 Рос АН, Сиб. отделение, Центр. Сиб. Бот. Сад. - Новосибирск, 1996. - 33с.
14. Определитель пресноводных водорослей СССР. Сине-зеленые водоросли / Полянский В.И. - М.: изд-во Советская наука, 1953. - Вып.2. - 560с.
15. Определитель пресноводных водорослей СССР. Эвгленовые водоросли / Попова Т.Г. - М.: изд-во Советская наука, 1955. - Вып.7 - 288с.
16. Оформление курсовых и дипломных работ: метод. рекомендации / сост.С.М. Григорьевская, Е.Ю. Кичигина, В.С. Крылова; Том. гос. ун-т, науч. б-ка, Библиогр. информ. центр. - Томск, 2007. - 39с.
17. Практикум по физиологии растений / под ред.Н. Н. Третьякова. - 2-е изд-е, перераб. и доп. - М.: Колос, 1982. - 271с.
18. Ресурсы поверхностных вод СССР. Т15. Алтай и Западная Сибирь. вып.2 Средняя Обь. Л.: Гидрометеоиздат, 1973. - 408с.
19. Романенко В.И. сравнение кислородного и радиоуглеродного методов определения интенсивности фотосинтеза фитопланктона / Романенко В.И. // Труды ин-та биологии внутренних вод АНСССР. Микрофлора, фитопланктон и высшая растительность внутренних водоемов. - 1967. - вып.15. - С.54-60
20. Савичев О.Г. Исследование эколого-геохимического состояния речных вод бассейна Средней Оби: диссертация на соискание ученой степени КГН.11.00.11. - Томск, 1996. - 217с.
21. Савичев О.Г. Реки Томской области: состояние, охрана и использование / О.Г. Савичев. - Томск: изд-во ТПУ, 2003. - 202с.
22. Смирнова А.Н. К вопросу о санитарно-показательном значении некоторых наиболее распространенных водорослей // Гидробиологический журнал. - 1965. - т.1. - №4. - С.53-59
23. Тополов А.А. Уточнение скляночного метода определения деструкции органического вещества в водоемах // Биология внутренних вод. - 1970. - №6. - С.12-15
24. Трухин Н.В. К определению скорости фотосинтеза водорослей кислородным методом // Труды ин-та биологии внутренних вод. Лучистые факторы жизни водных организмов. - 1967. - вып.14. - С.3-9
25. Шмидт В.М. Математические методы в ботанике / В.М. Шмидт - Л.: Изд-во Ленингр. ун-та, 1984. - 288с.
26. Шорникова Е.А. Характеристика гидрохимического режима водотоков широтного отрезка Средней Оби / Шорникова Е.А. // Водное хозяйство России. - 2007. - №2. - С.57-72
Приложения
Приложение А
Таблица 1 Доминирующие виды на правом берегу р. Обь у п. Половинка
| МЕСЯЦ | ПРОЦЕНТЫ | ДОМИНИРУЮЩИЕ ВИДЫ |
| Май |
23 23 21 15,8 |
Ankistrodesmus pseudomirabilis var. spiralis Korschik (Chlorophyta) Gonatozygon monotaenium De Bary var. monotaenium (Chlorophyta) Closterium acerosum (Schr) Ehr. f. acerosum (Chlorophyta) Pleurotaenium baculoides (Roy et Biss) Playf (Chlorophyta) |
| Июнь |
18,77 18,77 12,5 12,5 12,5 12,5 12,5 |
Closterium setaceum Ehr (Chlorophyta) Closterium kuetzingii Breb var. kuetzingii (Chlorophyta) Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta) Asterionella formosa Hass (Bacillariophytа) Fragillaria construens (Ehr) Grum var. Subsalina Hust (Bacillariophytа) Scenedesmus quadricada (Turp) Breb var. abundans Kirchn (Chlorophyta) Ankistrodesmus pseudomirabilis var. spiralis Korschik (Chlorophyta) |
| Июль |
47,36 10,5 10,5 13,6 11,3 12,49 |
Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta) Gonium pectorale Müll (Chlorophyta) Nostoc carneum Ag. (Cyanophyta) Palmella microspora Korschik (Chlorophyta) Phormidium frigidum F. E. Fritsch (Cyanophyta) Crucigenia tetrapedia (Kirhn) W. et. G West (Cyanophyta) |
| Август |
27,8 19,99 13,32 13,32 19,98 |
Nostoc carneum Ag. (Cyanophyta) Oscillatoria kisselevii Anissim (Cyanophyta) Anabaena knipowinschii Ussatsch (Cyanophyta) Scenedesmus bijugatus (Turp) Kütz var. bijugatus (Chlorophyta) Oscillatoria kisselevii Anissim (Cyanophyta) |
| Сентябрь |
13,69 13,69 34,7 21,6 19,88 12,92 |
Trachelomonas lacustris var. Sabutata (Skv) Popova (Euglenophyta) Palmella microspora Korschik (Chlorophyta) Anabaena vugieri Denis et fremy (Cyanophyta) Anabaena echinospora Skuja (Cyanophyta) Ankistrodesmus longissimus (Lemm) Wille var. acicularis (Chod) Brunnth (Chlorophyta) Ankistrodesmus acicularis (A. Br) Korschik (Chlorophyta) |
| ПРИЛОктябрь |
63,62 59,98 |
Microcystis pulverea (Wood) Forti emend f. parasitica (Kűtz) Elenk (Cyanophyta) Oscillatoria kisselevii Anissim (Cyanophyta) |
Приложение В
Таблица №2 Видовой состав и экологическая характеристика представителей
| виды | 2007год | 2008 год | рН | соленость | сапробность |
|
отдел Bacillariophyta |
|||||
|
класс Centrophyceae |
|||||
|
Coscinodiscaceae |
|||||
| 1. Melosira islandica O. Müll subsp. islandica Hust | - | 0,126/9 | |||
| 2. Melosira undulata Kűtz var. undulate | 0,189/5 | - | |||
|
класс Pennanophyceae |
|||||
|
порядок Araphales |
|||||
|
сем-во Tabellariaceae |
|||||
| 3. Tabellaria fenestrata Kűtz var intermedia Grun | 0,126/7 | - | β | ||
|
сем-во Fragillariaceae |
|||||
| 4. Asterionella formosa Hass | - | 0,126/5 | 0 | ||
| 5. Fragillaria construens (Ehr) Grum var. Subsalina Hust | - | 0,126/6 | hl | 0 | |
| 6. Meridion curculare Ag. var curculare | 0,063/9 | - | Х-0 | ||
| 7. Opephora martyi f. intermedia Komar. var. nov | - | 0,0989/9 | |||
| 8. Opephora martyi Herib var. martyi f. martyi | - | 0,1415/8 | |||
| 9. Synedra tabulata (Ag) Kütz tabulata | - | 0,063/6 | mh | 0 | |
|
порядок Raphales |
|||||
|
сем-во Eunotiaceae |
|||||
| 10. Eunotia bigibba Kütz var. pumila Grun | - | 0,126/8 | |||
|
сем-во Achnanthaceae |
|||||
| 11. Coccoineus disculus var. disculus (Schum) Cl | - | 0,063/8 | oh | ||
| сем-во Naviculaceae | |||||
| 12. Amphora ovalis var. gracilis Kütz | - | 0,0989/9 | acf | oh | - |
| 13. Amphora ovalis var. libyca Kütz | - | 0,0989/9 | |||
| 14. Amphora ovalis var. pediculus Kütz | - | 0,063/8 | 0 | ||
| 15. Cymbella cistula (Hemp) Grum var. Cistula | - | 0,063/7 | |||
| 16. Cymbella skvortzovii Skabitsch | - | 0,063/8 | |||
| 17. Cymbella prostrata (Berk) Cl | 0,189/5 | - | β | ||
| 18. Diploneis parma Cl | - | 0,063/7 | |||
| 19. Navicula amphibola Cl | - | 0,0989/9 | oh | ||
| 20. Navicula crucicula (W. Sm) Donk var. obtusata Grun | - | 0,063/8 | |||
| 21. Navicula jasnitskyi Skv et Mey | - | 0,063/9 | |||
| Navicula oblongata Kütz var. oblongata | - | 0,063/5 | i | ||
| 22. Navicula peregrina (Ehr) Kütz var. asiatica Skv. | - | 0,063/6 | |||
| 23. Navicula tuscula (Ehr) Grun | - | 0,063/7 | i | 0 | |
| 24. Neidum dilatatum var. chromica Komar. var. nov. | 0,063/10 | - | |||
| 25. Pinnularia bigibba Ehr. Var. mesogondyla f. interrupta | - | 0,063/6 | |||
| 26. Pinnularia lata (Breb) W. Sm. var. lata | - | 0,0989/9 | oh | ||
| 27. Pinnularia lata (Breb) W. Sm. var. minor Grun | - | 0,063/7 | |||
| сем-во Epithemiaceae | |||||
| 28. Denticula elegans Kütz | - | 0,0472/8 | oh | ||
| 29. Rhopalodia parallela (Grun) O. Müll | - | 0,0472/8 | oh | ||
| сем-во Nitzschiaceae | |||||
| 30. Nitzschia gracilis Hantzsch var. minor Skabitsch | - | 0,0472/8 | ind | i | 0 |
| 31. Nitzschia vermicularis (Kütz) Grun | - | 0,0472/8 | ind | i | |
| 32. Nitzschia hantzschiana Rabenh | 0,189/5 | - | |||
| 33. Nitzschia obtusa V. Sm. var. obtuse | 0,879/5 | - | 0 | ||
|
сем-во Surirellaceae |
|||||
| 34. Surirella didyma Kűtz | 0,063/5 | - | |||
| 35. Surirella robusta var. splendida Ehr | 0,252/10 | - | |||
|
отдел Chlorophyta |
|||||
|
порядок Volvocales |
|||||
|
сем-во Chlamydomonadaceae |
|||||
| 1. Chlamydomonas conferta Korschik | - | 0,063/7 | |||
| 2. Chlamydomonas macropirenoidosa Skuja | - | 0,126/7 | |||
|
сем-во Volvocaceae |
|||||
| 3. Gonium pectorale Müll | - | 0,566/7 | |||
| 4. Gonium sociale Warm | - | 0,063/7 | β | ||
|
порядок Tetrasporales |
|||||
|
сем-во Nautococcaceae |
|||||
| 5. Apiococcus consociatus Korschik | 0,252/9 | - | |||
|
порядок Chroococcales |
|||||
|
сем-во Clorococcaceae |
|||||
| 6. Tetraëdron caudatum var. longispinum Lemm | - | 0,063/7 | β | ||
| 7. Tetraëdron incus (Teil) G. M. Smith | - | 0,0472/7 | |||
| 8. Tetraëdron minutum (A. Br) Hansg f. minutum | - | 0,063/7 | β | ||
| 9. Tetraëdron minutissimum Korschik | - | 0,063/8 | |||
| 10. Dictiococcus mucosus Korschik | - | 0,063/7 | |||
| 11. Schroederia setigera (Schroed) Lemm | - | 0,1415/7 | |||
| 12. Korschikoviella gracilipes (Lambert) Silva (=Lambertia gracilipes (Lambert) Korschik) | - | 0,063/7 | |||
| 13. Korschikoviella limnetica (Lemm) Silva (=Lambertia limnetica (Lemm) Korschik) | - | 0,409/9 | |||
| 14. Korschikoviella spatulifera (Korschik) Silva | - | 0,047/9 | |||
| 15. Desmotractum indutum (Geitl) Pasch | - | 0,063/7 | |||
| 16. Ropalosolen cylindrica (Korschik) Fott | - | 0,047/10 | |||
|
сем-во Palmellaceae |
|||||
| 17. Palmella microspora Korschik | - | 0,377/7 | |||
| 18. Sphaerocystis planctonicus (Korschik) (=Palmellocystis planctonica Korschik) | - | 0,063/8 | |||
|
сем-во Oocystaceae |
|||||
| 19. Ankistrodesmus acicularis (A. Br) Korschik | - | 0,943/9 | β | ||
| 20. Ankistrodesmus braunii (Näg) Collius | - | 0,0943/9 | |||
| 21. Ankistrodesmus closteroides (Printz) Korschik | - | 0,0989/9 | |||
| 22. Ankistrodesmus longissimus (Lemm) Wille var. acicularis (Chod) Brunnth | - | 0,1415/7 | |||
| 23. Ankistrodesmus longissimus (Lemm) Wille var. longissimus | - | 0,236/9 | |||
| 24. Ankistrodesmus obtusus Korschik | - | 0,1886/8 | |||
| 25. Ankistrodesmus pseudomirabilis var. pseudomirabilis | - | 0,11/9 | |||
| 26. Ankistrodesmus pseudomirabilis var. spiralis Korschik | - | 0,189/5 | |||
| 27. Truebaria varia Tiff et Ahlstr | - | 0,063/6 | |||
| 28. Trochiscia aciculifera (Lagerh) Hansg | - | 0,063/8 | ind | ||
| 29. Hyaloraphidium contortum Pasch et Korschik var. contortum | - | 0,11/7 | |||
| 30. Hyaloraphidium contortum Pasch et Korschik var. tenuissimum Korschik | - | 0,063/7 | |||
| 31. Eremosphaera viridis De Bary | - | 0,063/9 | acf | ||
| 32. Kirchneriella intermedia var. major Korschik | - | 0,063/7 | |||
| 33. Oocystis lacustris Chod | - | 0,0472/7 | |||
| 34. Oocystis marsonii Lemm (=O. crassa Wittr) | - | 0,063/7 | |||
| 35. Oocystis pelagica Lemm | - | 0,063/8 | |||
| 36. Oocystis pusilla Hansg | - | 0,063/7 | |||
| 37. Oonephris obesa (West) Fott (=Oocystis gigas Archer. F. minor W. West, G. West) | - | 0,063/8 | |||
| 38. Nephrocytium agardhianum Näg | - | 0,063/7 | |||
|
сем-во Radiococcaceae |
|||||
| 39. Coenocystis subcilindrica Korschik | 0,314/5 | - | |||
|
сем-во Scenedesmaceae |
|||||
|
40. Actinastrum hantschii Lagerh var. gracile Roll |
0,377/6 | 0,189/7 | ind | i | β |
|
41. Actinastrum hantschii Lagerh var. hantschii |
0,063/7 | 0,063/7 | |||
| 42. Crucigenia quadrata Morr | 0,189/6 | - | |||
|
43. Crucigenia tetrapedia (Kirhn) W. et. G West |
0,063/8 | 0,126/9 | ind | i | β |
|
44. Scenedesmus acuminatus (Legerh) Chod var. acuminatus (=Sc. falcatus Chod) |
0,063/8 | 0,0943/8 | |||
| 45. Scenedesmus acuminatus (Legerh) Chod var. bernardii (Smith) Deduss | - | 0,063/7 | |||
| 46. Scenedesmus acuminatus (Legerh) Chod var. biseriatus Reinh | - | 0,063/9 | β | ||
|
47. Scenedesmus apiculatus (W. West, G. West) |
0,189/6 | 0,0472/8 | i | ||
| 48. Scenedesmus bijugatus (Turp) Kütz var. bijugatus | 0,252/9 | 0,0943/8 | |||
| 49. Scenedesmus bijugatus var. octacellularis (=Sc. Octacellularis Kisselev) | - | 0,063/7 | |||
| 50. Scenedesmus obliquus (Turp) Kütz var. alternans Christ | - | 0,0472/7 | |||
| 51. Scenedesmus quadricada (Turp) Breb var. abundans Kirchn | - | 0,126/6 | |||
| 52. Scenedesmus quadricada (Turp) Breb var. lefevrii (Delf) Deduss | - | 0,0472/7 | |||
| 53. Tetradesmus cumbricus var. apiculatus Korschik | - | 0,063/8 | |||
| 54. Tetrastum glabrum (Roll) Ahlstr et Tiff | - | 0,063/7 | |||
| 55. Tetrastum punctatum (Schm) Ahlstr et Tiff | - | 0,063/6 | |||
| 56. Tetrachlorella alternans Korschik | - | 0,0472/7 | |||
| сем-во Hydrodictyaceae | |||||
| 57. Pediastrum boryanum (Turp) Menegh | - | 0,0472/8 | ind | i | |
| 58. Pediastrum simplex Meyen | 0,063/9 | 0,063/7 | ind | i | β |
| 59. Pediastrum dulex Meyen var. clathratum (A. Br) Lagerh | - | 0,063/5 | ind | i | β |
| 60. Pediastrum dulex Meyen var. inflata Wolosz | 0,063/8 | - | |||
| 61. Pediastrum dulex Meyen var. setigera (Corda) Rabenh | - | 0,063/7 | |||
| 62. Pediastrum tetrapodum Mor. - Wod. | - | 0,189/8 | |||
| 63. Pediastrum tetras var. tetras | - | 0,063/8 | ind | i | β |
| порядок Ulothricales | |||||
| сем-во Ulothricaceae | |||||
| 64. Ulotrix subtilissima Rabenh | - | 0,0989/9 | |||
| 65. Ulotrix variabilis Kűtz | - | 0,0472/7 | |||
|
66. Ulotrix zonata Kűtz |
0,063/10 | 0,0472/8 | ind | i | 0 |
|
порядок Zygnematales |
|||||
|
сем-во Zygnemataceae |
|||||
| 67. Mougeotia laetevirens Wittr | - | 0,063/7 | |||
| 68. Spirogyra mirabilis (Hass) Kűtz | - | 0,063/5 | |||
|
сем-во Mesotaeniaceae |
|||||
| 69. Spirotaenia condensata Breb | - | 0,063/7 | |||
| 70. Gonatozygon monotaenium De Bary var. monotaenium | - | 0,189/5 | |||
| 71. Gonatozygon monotaenium De Bary var. pilosellum Nordst | - | 0,173/9 | |||
|
сем-во Desmidiaceae |
|||||
| 72. Closterium acerosum (Schr) Ehr. f. acerosum | - | 0,252/5 | ind | i | α |
| 73. Closterium acerosum (Schr) Ehr. f. elongatum (Breb) Kossinsk | - | 0,2515/9 | |||
| 74. Closterium acerosum (Schr) Ehr. f. minus (Hantzsch) Kossinsk | - | 0,063/8 | |||
| 75. Closterium acutum (Lyngb) Breb | - | 0,063/9 | |||
| 76. Closterium calosporum var. calosporum Wittr | - | 0,063/8 | acf | ||
| 77. Closterium gracile Breb f. gracile | - | 0,063/7 | |||
| 78. Closterium kuetzingii Breb var. kuetzingii | - | 0,189/6 | ind | ||
| 79. Closterium lanceolanum Kűtz | - | 0,409/9 | |||
| 80. Closterium littorale Gay f. littorale | - | 0,063/9 | |||
|
81. Closterium manschuricum Skvortz |
0,126/10 | 0,11/9 | |||
| 82. Closterium maculentum Breb | - | 0,063/9 | acf | ||
| 83. Closterium moniliferum (Bory) Ehr. concavum Klebs | - | 0,063/9 | i | β | |
| 84. Closterium parvulum Näg f. majus W. West | - | 0,0989/9 | i | β | |
| 85. Closterium peracerosum var. elegans W. West | - | 0,0472/7 | |||
| 86. Closterium peracerosum var. peracerosum Gay | - | 0,063/5 | |||
| 87. Closterium praelеugum Breb | - | 0,063/9 | |||
| 88. Closterium rostratum Ehr. var. brevirostratum (W. West) Kossinsk | - | 0,063/5 | |||
| 89. Closterium setaceum Ehr | - | 0,252/7 | acf | ||
| 90. Closterium siliqua W. West, G. West | - | 0,063/7 | |||
| 91. Closterium tumidum Johns | - | 0,063/8 | acf | ||
| 92. Closterium regulare Breb. | 0,063/5 | - | |||
| 93. Closterium littorale Gay f. minus Komarenko | 0,063/10 | - | |||
| 94. Cosmarium botrytis Menegh var. botrytis | - | 0,063/7 | ind | i | α |
| 95. Cosmarium ochthodes Nordst | - | 0,063/7 | |||
| 96. Cosmarium subcostatum Nordst | - | 0,0472/7 | |||
| 97. Cosmarium turgidum Breb | - | 0,0472/7 | |||
| 98. Docidium baculum Breb | - | 0,063/9 | acf | ||
| 99. Penium spirostriolatum Barker | 0,063/7 | - | |||
|
100. Pandorina morum (Müll) Bory |
0,063/8 | 0,063/9 | ind | i | β |
| 101. Pleurotaenium baculoides (Roy et Biss) Playf | - | 0,189/5 | acf | ||
| 102. Pleurotaenium coronatum (Breb) Rabenh var. nodulosum W. West | - | 0,063/5 | |||
| 103. Pleurotaenium ehrenbergii (Breb) De Bary | - | 0,063/5 | |||
| 104. Pleurotaenium minutum (Ralfs) Delf var. minutum | 0,063/5 | 0,047/9 | |||
| 105. Pleurotaenium minutum (Ralfs) Delf var. ractissimum (W. West, G. West) W. Krieg | - | 0,063/6 | |||
| 106. Pleurotaenium trabecula (Ehr) Näg f. trabecula | - | 0,063/5 | |||
| 107. Staurastum dickei Ralfs f. isthmolatissimo Wille | - | 0,063/7 | acf | ||
| 108. Staurastum cuspidatum Breb | - | 0,126/7 | |||
| 109. Staurastum oxyacanthum Arch var. polyacanthum Nordst | - | 0,0472/7 | |||
| 110. Staurastum punctulatum var. pygmaemum (Breb) W. West, G. West | - | 0,063/7 | |||
| отдел Cyanophyta | |||||
| класс Chroococcophyceae | |||||
| порядок Chroococcales | |||||
| сем-во Coccobactreaceae | |||||
| 1. Dactyllococcopsis planctonica Teiting | - | 0,063/7 | |||
| 2. Dactyllococcopsis rhaphidiodes f. falciformes Printz | - | 0,0989/9 | |||
| 3. Dactyllococcopsis scenedesmoides Nyg | - | 0,0472/7 | |||
| 4. Synechoococcus elongates Näg | - | 0,0472/7 | |||
| 5. Synechocystis Pevalekii Erceg | - | 0,0472/8 | |||
| 6. Rhabdoderma lineare Schmidle et Laut emend Hollerb f. lineare | - | 0,063/7 | |||
| сем-во Merismopediaceae | |||||
| 7. Merismopedia major (Smith) Geitl | - | 0,0472/7 | |||
| 8. Merismopedia tenuissima Lemm | - | 0,0472/8 | hl | β-α | |
| сем-во Microcystidaceae | |||||
| 9. Aphanothece clathrata W. et G. S. West | - | 0,063/7 | |||
| 10. Aphanothece elabens (Breb) Elenk | - | 0,0472/7 | |||
| 11. Microcystis aeruginosa Kűtz emend Elenk f. flos-aquae (Wittr) Elenk | - | 0,063/7 | hl | ||
| 12. Microcystis aeruginosa Kűtz emend Elenk f. marginata (Megenh) (=Microcystis marginata (Megenh) Kűtz) | - | 0,3459/9 | |||
| 13. Microcystis pulverea (Wood) Forti emend f. parasitica (Kűtz) Elenk | - | 0,33/10 | i | β | |
|
сем-во Gloeocapsaceae |
|||||
| 14. Gloeocapsa limnetica (Lemm) Hollerb | - | 0,0943/7 | i | ||
| 15. Gloeocapsa minuta (Kűtz) Hollerb ampl | - | 0,063/8 | ind | hl | 0 |
| 16. Gloeothece rupestris Nag Born (Lyngb) | 0,377/10 | - | |||
|
сем-во Coleosphaeriaceae |
|||||
| 17. Marssoniella elegans Lemm (=Coelesphaerium radiatum G. M. Smith) | - | 0,063/7 | |||
|
класс Chamaesiphonophyceae |
|||||
|
порядок Pleurocapsales |
|||||
|
сем-во Pleurocapsaceae |
|||||
| 18. Pleurocapsa minor Hansg emend Crietf | - | 0,063/7 | |||
| 19. Chroococcopsis gigantea Cleitl | - | 0,063/7 | |||
|
порядок Dermocarpales |
|||||
|
сем-во Chamaesiphonaceae |
|||||
| 20. Chamaesiphon curvatus (Borzi) Nordst | - | 0,0989/9 | |||
|
класс Hormogoniphyceae |
|||||
|
порядок Stigonematales |
|||||
|
сем-во Stigonemataceae |
|||||
| 21. Stigonema ocellatum (Dillw) Thur. sensulat Elenk | - | 0,063/7 | acf | ||
|
порядок Nostocales |
|||||
|
сем-во Nostocaceae |
|||||
| 22. Nostoc carneum Ag. | - | 0,63/8 | |||
| ем-во Anabaenaceae | |||||
| 23. Anabaena affinis Lemm | - | 0,0472/7 | |||
| 24. Anabaena echinospora Skuja | 0,566/7 | 0,314/9 | |||
| 25. Anabaena hallensis (Janez) Born of Flas | 0,314/9 | ||||
| 25. Anabaena hallensis (Janez) Born of Flas | |||||
| 26. Anabaena Hassalii (Kűtz) Wittr. | 0,063/8 | 0,063/7 | |||
| 27. Anabaena heterospora Nygaard | - | 0,189/7 | |||
| 28. Anabaena knipowinschii Ussatsch | - | 0,1415/7 | |||
| 29. Anabaena oscillaroides f. torulosa (Lagerh) Elenk | - | 0,063/7 | hl | ||
| 30. Anabaena planctonica Brunnth | - | 0,0472/8 | |||
| 31. Anabaena sibirica (Popova et Degt) Elenk | - | 0,189/7 | |||
| 32. Anabaena solitaria Kleb f. zinserlingii (Kossinsk) Elenk | - | 0,0472/8 | |||
| 33. Anabaena spiroides Kleb. | 0,44/7 | 0,189/7 | 0-β | ||
| 34. Anabaena tenericautis (Nygaard) Kossinsk f. longispora | - | 0,063/7 | |||
| 35. Anabaena variabilis Kűtz rotundospora Hollerb | - | 0,0472/7 | |||
| 36. Anabaena vugieri Denis et fremy | - | 0,503/9 | |||
| 37. Anabaena Sheremetievi Elenk | 0,692/7 | - | |||
| 38. Abaenopsis Arnoldii Aptek | - | 0,063/7 | |||
| сем-во Nodulariaceae | |||||
| 39. Aulosira laxa Kirhn | 0,063/7 | - | |||
| сем-во Scytonemataceae | |||||
| 40. Scytonema drilosiphon Elenk et. V. Pljansk | 0,063/5 | - | |||
| сем-во Rivulariaceae | |||||
| 41. Gloeotrichia kamschatica (Elenk) V. Poljansk | - | 0,0472/8 | |||
| 42. Rivularia planctonica Elenk | 0,063/6 | - | |||
| порядок Oscillatoriales | |||||
| сем-во Pseudonostocaceae | |||||
| 43. Pseudoanabaena galeata Eöcher | 0,063/7 | - | |||
| сем-во Oscillatoriaceae | |||||
| 44. Lyngbia martensiana Menegh | - | 0,0472/8 | |||
| 45. Oscillatoria kisselevii Anissim | - | 0,1886/8 | hl | ||
| 46. Oscillatoria lacustris (Kleb) Geitl | - | 0,0472/8 | i | ||
| 47. Oscillatoria minima Gickhl | - | 0,2358/7 | |||
| 48. Oscillatoria mirabilis Böcher | - | 0,063/9 | |||
| 49. Oscillatoria prolifica (Grev) Gom | - | 0,063/7 | |||
| 50. Oscillatoria Shultzii Lemm | - | 0,063/7 | |||
| 51. Oscillatoria splendida Grev | - | 0,0472/8 | α | ||
| 52. Phormidium ambiguum Gom of novae-semliae (Schirsch) Elenk | - | 0,063/5 | ind | i | |
| 53. Phormidium curtum Hollerb | 0,063/5 | - | |||
| 54. Phormidium frigidum F. E. Fritsch | - | 0,314/7 | |||
| 55. Phormidium mucola Hub. - Pestalosii et Naum | - | 0,063/7 | |||
| 56. Romeria elegans (Wolosz) Koszw | - | 0,063/7 | |||
| сем-во Schizothricaceae | |||||
| 57. Schizothrix friesii (Ag) Com | - | 0,0472/8 | |||
| отдел Euglenophyta | |||||
| порядок Euglenales | |||||
| сем-во Euglenaceae | |||||
| 1. Euglena granulata (Klebs) Schmitz | - | 0,0472/8 | oh | 0-β | |
| 2. Euglena hemichromata Skuja | - | 0,063/8 | |||
| 3. Euglena satelles Brasl. - Spect | - | 0,063/7 | |||
| 4. Euglena texta (Duj) Hübner | 0,063/6 | - | 0 | ||
| 5. Euglena mucifera Mainx | 0,063/5 | - | |||
| 6. Euglena acus Ehr. | 0,063/6 | - | β | ||
| 7. Euglena acus var. hyalinа | 0,063/8 | - | |||
| 8. Lepocinclis fusiformes (Carter) Lemm | - | 0,063/8 | hl | 0-β | |
| 9. Strombomonas Deflandrei (Roll) Delf | 0,063/8 | - | |||
| 10. Strombomonas samarae (Swir) Popova | - | 0,063/7 | |||
| 11. Trachelomonas conica Playf. (=Tr. conica Playf f. Punctata Delf) | - | 0,063/7 | |||
| 12. Trachelomonas lacustris Drez. Emend Balech | - | 0,063/8 | acf | ||
| 13. Trachelomonas lacustris var. Sabutata (Skv) Popova | - | 0,189/8 | acf | ||
| 14. Trachelomonas Matvienkovi Popova | - | 0,0472/8 | acf | ||
| 15. Trachelomonas mirabilis Swir var. minor Woronich | - | 0,063/10 | acf | ||
| 16. Trachelomonas intermedia Dang | 0,126/9 | - | 0 | ||
| 17. Trachelomonas armata (Ehr) Stein | 0,063/6 | - | |||
|
сем-во Astasiaceae |
|||||
| 18. Astasia inflata f. fusiformes (Skuja) Popova (=Astasia inflata forma Skuja) | - | 0,063/7 | acf | ||
|
отдел Chrysophyta |
|||||
|
сем-во Euchromulinaceae |
|||||
| 1. Chromulina pyrum Parsch | 0,063/10 | - | |||
| 2. Chrysoglena verricosa Wisl | 0,063/10 | - |
Приложение С
Таблица №3 Единично встреченные виды микроводорослей (Средняя Обь, п. Половинка).
| месяц | процент | виды |
| май | - | нет |
| июнь | - | нет |
| июль |
2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 2,27 |
Mougeotia laetevirens Wittr (Chlorophyta) Aphanothece clathrata W. et G. S. West (Cyanophyta) Closterium gracile Breb f. gracile (Chlorophyta) Closterium setaceum Ehr (Chlorophyta) Crucigenia tetrapedia (Kirhn) W. et. G West (Chlorophyta) Pediastrum simplex Meyen (Chlorophyta) Tetrastum punctatum (Schm) Ahlstr et Tiff (Chlorophyta) Desmotractum indutum (Geitl) Pasch (Chlorophyta) Phormidium mucola Hub. - Pestalosii et Naum (Cyanophyta) Spirotaenium condensata Breb (Chlorophyta) Navicula tuscula (Ehr) Grun (Bacillariophyta) Oscillatoria Shultzii Lemm (Cyanophyta) Microcystis aeruginosa Kűtz emend Elenk f. flos-aquae (Wittr) Elenk (Cyanophyta) Romeria elegans (Wolosz) Koszw (Cyanophyta) |
| август |
2,77 2,77 2,77 2,77 2,77 2,77 2,77 2,77 2,77 2,77 |
Lepocinclis fusiformes (Carter) Lemm (Euglenophyta) Sphaerocystis planctonicus (Korschik) (=Palmellocystis planctonica Korschik) (Chlorophyta) Euglena hemichromata Skuja (Euglenophyta) Closterium acerosum (Schr) Ehr. f. minus (Hantzsch) Kossinsk (Chlorophyta) Oocystis pelagica Lemm (Chlorophyta) Oonephris obesa (West) Fott (=Oocystis gigas Archer. F. minor W. West, G. West) (Chlorophyta) Navicula crucicula (W. Sm) Donk var. obtusata Grun (Bacillariophyta) Tetraëdron minutissimum Korschik (Chlorophyta) Cymbella skvortzovii Skabitsch (Bacillariophyta) Closterium calosporum var. calosporum Wittr (Chlorophyta) |
| август |
2,77 2,77 2,77 2,77 |
Closterium setaceum Ehr (Chlorophyta) Closterium tumidum Johns (Chlorophyta) Scenedesmus bijugatus (Turp) Kütz var. bijugatus (Chlorophyta) Truebaria varia Tiff et Ahlstr (Chlorophyta) |
| сентябрь |
4,56 4,65 4,65 4,56 4,56 4,56 4,56 4,56 4,56 4,56 4,56 4,56 4,34 4,34 4,34 4,34 4,34 4,34 3,64 4,97 1,99 2,32 |
Coccoineus disculus var. disculus (Schum) Cl (Bacillariophyta) Trachelomonas lacustris Drez. Emend Balech (Euglenophyta) Closterium tumidum Johns (Chlorophyta) Tetradesmus cumbricus var. apiculatus Korschik (Chlorophyta) Scenedesmus bijugatus (Turp) Kütz var. bijugatus (Chlorophyta) Pediastrum tetras var. tetras (Chlorophyta) Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta) Trochiscia aciculifera (Lagerh) Hansg (Chlorophyta) Gloeocapsa minuta (Kűtz) Hollerb ampl (Cyanophyta) Amphora ovalis var. pediculus Kütz (Bacillariophyta) Pandorina morum (Müll) Bory (Chlorophyta) Scenedesmus acuminatus (Legerh) Chod var. biseriatus Reinh (Chlorophyta) Anabaena spiroides Kleb (Cyanophyta) Actinastrum hantschii Lagerh var. gracile Roll (Chlorophyta) Closterium maculentum Breb (Chlorophyta) Eremosphaera viridis De Bary (Chlorophyta) Scenedesmus acuminatus (Legerh) Chod var. acuminatus (=Sc. falcatus Chod) (Chlorophyta) Microcystis aeruginosa Kűtz emend Elenk f. flos-aquae (Wittr) Elenk (Cyanophyta) Gonatozygon monotaenium De Bary var. pilosellum Nordst (Chlorophyta) Ankistrodesmus longissimus (Lemm) Wille var. longissimus (Chlorophyta) Ankistrodesmus braunii (Näg) Collius (Chlorophyta) Ankistrodesmus pseudomirabilis var. pseudomirabilis (Chlorophyta) |
| сентябрь |
1,33 2,65 1,99 1,99 1,99 1,99 1,99 1,99 1,99 1,99 1,99 1,99 1,33 1,33 1,33 1,33 1,33 1,33 1,33 1,33 0,99 0,99 0,99 0,99 |
Closterium manschuricum Skvortz (Chlorophyta) Schroederia setigera (Schroed) Lemm (Chlorophyta) Amphora ovalis var. gracilis Kütz (Bacillariophyta) Dactyllococcopsis rhaphidiodes f. falciformes Printz (Cyanophyta) Opephora martyi f. intermedia Komar. var. nov (Bacillariophyta) Ulotrix subtilissima Rabenh (Chlorophyta) Chamaesiphon curvatus (Borzi) Nordst (Chlorophyta) Navicula amphibola Cl (Bacillariophyta) Pinnularia lata (Breb) W. Sm. var. lata (Bacillariophyta) Ankistrodesmus closteroides (Printz) Korschik (Chlorophyta) Amphora ovalis var. libyca Kütz (Bacillariophyta) Closterium parvulum Näg f. majus W. West (Chlorophyta) Closterium praelcugum Breb (Chlorophyta) Docidium baculum Breb (Chlorophyta) Closterium tumidum Johns (Chlorophyta) Closterium acutum (Lyngb) Breb (Chlorophyta) Closterium littorale Gay f. littorale (Chlorophyta) Navicula jasnitskyi Skv et Mey (Bacillariophyta) Closterium moniliferum (Bory) Ehr. concavum Klebs (Chlorophyta) Oscillatoria mirabilis Böcher (Cyanophyta) Scenedesmus apiculatus (W. West, G. West) (Chlorophyta) Pleurotaenium minutum (Ralfs) Delf var. minutum (Chlorophyta) Korschikoviella spatulifera (Korschik) Silva (Chlorophyta) Pediastrum tetrapodus Mor. - Wod (Chlorophyta) |
| октябрь | нет |