Реферат: Функционирование нервных связей в зрительной системе
Реферат
На тему
Функционирование нервных связей в зрительной системе
Необходимые условия для поддержания функционирования нервных связей в зрительной системе
Уменьшение активности в зрительных путях является основным фактором, который стремится нарушить нормальное функционирование корковых нейронов. Однако клетки коры активируются не в ответ на диффузное освещение, также вызывающее активность, а на определенные контуры и формы. Для поддержания функционирования необходимы дополнительные факторы, гораздо более «тонкие» по своей природе. В частности, должно происходить взаимодействие между сигналами обоих глаз.
Бинокулярная депривация и роль конкуренции
Первым доводом против того, что уменьшение зрительной активности может привести к изменениям в работе нейронов, являются данные эксперимента, когда у детеныша обезьяны, новорожденного или рожденного путем кесарева сечения, закрывали сразу оба глаза. Исходя из предыдущего обсуждения, можно сделать предположение, что клетки в коре не будут управляться ни одним глазом. Однако, к удивлению, после бинокулярной депривации на протяжении 17 дней и дольше большинство корковых клеток выглядело полностью нормальными. Колонки ориентации имели строение, близкое к таковому у контрольной группы (рис. 1). Основным нарушением было то, что значительная часть клеток не получала бинокулярного управления (рис. 1). Кроме того, некоторые спонтанно активные клетки вообще не получали никакого управления, другим же для активности не требовался определенным образом ориентированный стимул. Тем не менее зоны коры, контролируемые обоими глазами, оставались равными в размерах, имея паттерн, аналогичный тому, что наблюдается у нормальных взрослых обезьян: в слое 4 клетки управлялись только одним глазом, четко определялись колонки при использовании авторадиографии или окраски на цитохромоксидазу. Бинокулярная депривация у котят приводила к сходным эффектам, за исключением того, что количество клеток, получающих бинокулярное управление, оставалось большим. При этом также не происходило уменьшение ветвистости аксонов ЛКТ, направляющихся в слой 4 коры. В то же самое время клетки всех слоев в пределах ЛКТ подвергались атрофии (уменьшению в размерах примерно на 40 %).
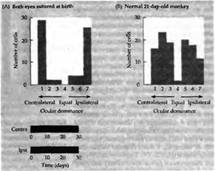
Рис. 1 Колонки зрительного доминирования после закрытия обоих глаз с рождения. (А) Обезьяна, на которой проводились исследования, была рождена посредством кесарева сечения, после чего веки обоих глаз были тотчас зашиты. Результаты были получены в возрасте 30 дней. Каждый из двух депривированных глаз все еще мог управлять клетками в зрительной коре. Рецептивные поля выглядели нормально, за исключением того, что некоторые клетки получали управление одновременно отобоих глаз. Время закрытия век показано внизу гистограммы. (В) Гистограмма глазного доминирования у нормальной обезьяны в возрасте 21 дня.
Выводом из этих экспериментов является то, что некоторые, но не все негативные эффекты, вызванные закрытием одного глаза, уменьшались или предотвращались при закрытии обоих глаз. Таким образом, импульсы от обоих глаз являются как бы конкурирующими за представление в корковых клетках, и эта конкуренция нарушается при закрытии одного глаза из двух.
Эффекты страбизма (косоглазия)
Аномалии, описанные ранее, были вызваны зашиванием век или использованием прозрачных диффузоров, приводящих к потере предметного зрения. Исходя из данных о том, что у детей, страдающих косоглазием или страбизмом (cross-eyed, strabismus, squint), а также имеющих аномальное расположение глаз (wall-eyed), также развивается слепота, Хьюбель и Визель вызывали искусственное косоглазие у котят и новорожденных обезьян, при помощи подрезания глазных мышц. В результате оптическая ось глаза отклонялась от нормальной. При этом освещение и паттерны стимуляции каждого глаза оставались без изменений.
Сначала результаты подобных опытов разочаровывали: после нескольких месяцев зрение в обоих глазах оперированного животного оставалось нормальным, и Хьюбель и Визелъ уже были готовы прекратить эту напряженную серию экспериментов (личное сообщение). Тем не менее они смогли получить следующие данные о работе корковых клеток. Отдельные корковые клетки имели нормальные рецептивные поля и живо отвечали на определенным образом ориентированные стимулы. Однако почти каждая клетка отвечала только на стимуляцию одного глаза, некоторые управлялись правым, некоторые — левым, но практически ни одна — обоими глазами. Клетки, как обычно, были сгруппированы в колонки по отношению к глазному предпочтению и к ориентации оси зрительного поля. Как и ожидалось, возникала атрофия в области ЛКТ, хотя колончатая архитектура слоя 4 оставалась без изменений. Почти полное отсутствие бинокулярного представления в коре показано на гистограмме, полученной от обезьяны с искусственно вызванным косоглазием (рис. 2). Критический период для смещенного (из-за косоглазия) восприятия, в течение которого возникают данные изменения, близок к критическому периоду монокулярной депривации.
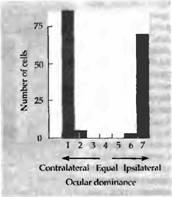
Рис. 2. Влияние косоглазия на формирование зрительного доминирования у обезьян. Глаза этой 3-летней обезьяны направлены в разные стороны, так как ей в возрасте 3 недель была проведена операция по подрезанию глазных мышц. Клетки коры получают управление либо от одного глаза, либо от другого, но не от обоих. Колонки глазного доминирования в слое 4 выглядят нормально для животного такого возраста.
Эти эксперименты представляют собой пример, когда все обычные параметры освещения в норме: количество попадающего в глаз света и паттерн стимуляции. Единственным отличием от нормы является то, что свет не падает на корреспондирующие регионы двух сетчаток. Фактором, который, скорее всего, приводит к потере бинокулярной конвергенции проекций отростков ЛКТ в кору, является недостаток соответствия в импульсах от обоих глаз. То есть гомологичные рецептивные поля обоих глаз должны быть скоординированы, чтобы возбуждение происходило одновременно. Последующие эксперименты говорят в пользу такой идеи.
В течение первых трех месяцев жизни глаза котенка были закрыты непрозрачной пластиковой пластиной, которая перемещалась с одного глаза на другой каждый день таким образом, чтобы два глаза получили одинаковый зрительный опыт, но в разное время. Результат был аналогичен данным экспериментов с искусственным косоглазием: клетки в основном управлялись либо правым, либо левым глазом, но не обоими. Поддержание нормальной бинокулярности зрения, таким образом, зависит не только от количества импульсов, но также и от надлежащего пространственного и временного соответствия активности в различных идущих к коре волокнах.
Изменения в ориентационном предпочтении
Логичным продолжением является вопрос, изменится ли ориентационное предпочтение кортикальных нейронов у животных, находящихся в условиях, когда они видят образ только одной ориентации. Экспериментальный подход, включающий в себя также конкуренцию между глазами и депривацию, был использован Карлсоном, Хьюбелем и Визелем. У новорожденной обезьяны были зашиты веки одного глаза. Животное содержалось в темноте, за исключением того времени, когда его голова помещалась в специальный держатель. В этом случае его голова фиксировалась строго вертикально и животное могло видеть вертикальные полоски оставшимся недепривированным глазом. Так как обезьяне всегда давали апельсиновый сок, когда она правильно устанавливала свою голову в держатель, она научилась быстро производить этот маневр. Таким образом, во время критического периода один глаз не получал никакой зрительной информации, а другой
видел только вертикальные полоски. После приобретения 57-и часов опыта в интервале между 12-м и 54-м днем с момента рождения регистрировался нормальный уровень корковой активности, когда клетки всех ориентации были организованы как обычно, в виде колонок. Как и ожидалось, наблюдалось доминирование недепривированного (открытого) глаза.
При проведении тестов для определения ориентационного предпочтения были получены результаты, показанные на рис. 2. И. Оба глаза одинаково отвечали на горизонтально ориентированные линии, однако на вертикальные гораздо эффективнее отвечал левый (открытый) глаз. Возможным объяснением этого факта может служить то, что ни один глаз не видел горизонтальные полоски или контуры на протяжении критического периода. Следовательно, стимуляция горизонтальными линиями является для них аналогом бинокулярной депривации, то есть конкуренция по отношению к ним была равной. Что касается вертикальных полосок, здесь открытый глаз получил гораздо больше опыта, по сравнению с закрытым, и «захватил» часть клеток в колонках вертикальной ориентации, которые первоначально принадлежали депривированному глазу. Подобные же результаты были получены Бонхоеффером с коллегами на котятах, которым представлялись только полоски определенной ориентации.
Критические периоды в развитии зрительной системы человека и их клиническое значение
Чувствительность котят и детенышей обезьян в ранние периоды жизни напоминает клинические наблюдения, выполненные на людях. Давно известно, что удаление непрозрачного хрусталика («катаракта») может привести к восстановлению зрения, даже если пациент был слепым на протяжении многих лет. И наоборот, катаракта может развиться у новорожденных или неполовозрелых детей, что часто приводит к слепоте. До экспериментов Хьюбеля и Визеля катаракты маленьким детям удаляли в позднем возрасте, когда они считались готовыми для этой операции. В результате формировалась перманентная слепота, без возможности к восстановлению. В настоящее время катаракты у маленьких детей удаляются хирургическим способом как можно раньше, в критические периоды, что обеспечивает очень благоприятный прогноз восстановления зрения.
Вторым клиническим применением полученных данных может служить известная медицинская манипуляция, используемая в прошлом для лечения детей с косоглазием и аномальным расположением глаз. Обычно на глаз, который видит хорошо, на долгий период ставится окклюзия, чтобы таким образом ребенок использовал более слабый глаз и разрабатывал его. Однако имеются данные, что это может привести к снижению остроты зрения депривированного глаза, в зависимости от возраста ребенка и времени окклюзии. Такая продолжительная окклюзия больше не используется в обычной медицинской практике. Клинические данные показывают, что наибольшей чувствительностью обладают дети первого года жизни, однако критический период может длиться и на протяжении нескольких лет.
Интересным является тот факт, что в глазодоминантных колонках, выявленных при помоши окраски на цитохромоксидазу слоя 4 зрительной коры у пациентов postmortem, обнаруживаются последствия монокулярной депривации в виде нарушений организации глазодоминантных колонок, подобно тому как это происходит у обезьян и котят. Хортон и Хокинг изучали паттерны окрашивания в области первичной зрительной коры у детей, которые в возрасте 1 недели перенесли хирургическое удаление одного глаза из-за опухоли. После смерти в их головном мозге, как и ожидалось, окрашивание в пределах слоя 4С было однородным, вместо отдельной территории для каждого глаза, как это наблюдается у нормальных людей и у пациентов, перенесших удаление глаза во взрослом состоянии. Как и ожидалось, длительное (с двухлетнего возраста) косоглазие у пациента не выявило изменений в ширине колонок, как было обнаружено после смерти в возрасте 79 лет.
Третьим примером клинической важности подобных экспериментов является удлинение (elongation) глазного яблока, которое происходило при сшивании век у обезьян в неонатальном периоде. Подобное удлинение приводило к размытости изображения и к близорукости (миопии). Известно, что у детей также развивается близорукость, если веки мешают зрению или если нарушается прозрачность роговицы. Хотя есть некоторые доказательства того, что определенную роль в этом играют нейромедиаторы и активность клеток, тем не менее механизмы удлинения глаза при закрытии век до сих пор остаются неизвестными.
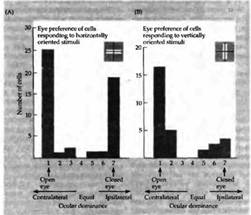
Рис. 3. Ориентационные предпочтения кортикальных клеток обезьяны с модифицированным зрительным опытом. Обезьяна содержалась в темной комнате. В возрасте 12 дней был закрыт правый глаз. Когда обезьяна правильно устанавливала свою голову в специальный держатель, она получала апельсиновый сок. При этом она видела своим левым глазом вертикальные полоски (специальный держатель для головы обеспечивал ее ровное положение). На протяжении в общей сложности 57 часов в возрасте между 12 и 54 днями один глаз видел только вертикальные линии, а другой не видел ничего. (А) Когда на экран проецировались горизонтально ориентированные световые стимулы, кортикальные клетки, управляемые как правым, так и левым глазом, отвечали одинаково хорошо. По этой диаграмме нельзя выявить признаков депривации, за исключением отсутствия бинокулярных клеток. (В) При использовании вертикально ориентированных стимулов левый глаз, который постоянно был открытым, управлял корковыми клетками более эффективно, чем правый. Диаграмма соответствует таковой после монокулярной депривации. Результаты позволяют предположить, что конкуренция за горизонтальные стимулы, которые не видел ни один глаз, была одинакова, а для вертикальных стимулов — различна (здесь преимущества имел левый, открытый глаз).
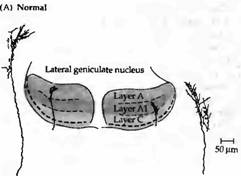
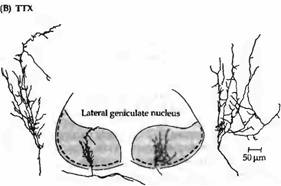
Рис. 4. Эффект подавления электрической активности тетродотоксином на ветвистость нервных волокон глазного нерва, заканчивающихся в ЛКТ. (А) У нормального котенка окончания зрительного нерва, помеченные при помощи пероксидазы хрена, ограничены пределом того слоя, где они в норме заканчиваются. (В) После применения тетродотоксина на протяжении 16 дней в эмбриональном возрасте, помеченные аксоны имеют гораздо большую ветвистость и не ограничены всего лишь одним слоем.
Литература
1. Adams, R. D., Victor, M., and Ropper, A. H. 1997. Principles of Neurology, 6th Ed. McGraw-Hill, New York.
2. Harlow, J. M. 1868. Publ. Mass. Med. Soc. 2: 328-334.
3. Driver, J., and Halligan, P. W. 199). Cogn. Neuropsy-chol. 8: 475-496.
4. Raff, M. 1996. Science 274: 1063.
5. Krauthammer, C. 2000. Time Magazine February 14: 76.