Реферат: Энергия в экологических системах. Принципы и концепции. Метрология и размер особей
ГОСУДАРСТВЕННО ОБРОЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРОЗОВАНИЯ
КАЗАНСКОГО ГОСУДАРСТВЕННОГО ТЕХНИЧЕСКОГО УНИВЕРСИТЕТА им.Туполева
КАФЕДРА ОБЩЕЙ ХИМИИ И ЭКОЛОГИИ
РЕФЕРАТ ПО КУРСУ ОБЩЕЙ ЭКОЛОГИИ.
Энергия в экологических системах. Принципы и концепции.Метаболизм и размер особей
Выполнил студент
Группы: 3212
Бурганов Д. А.
Проверил: доцент Гоголь Э. В.
Казань 2011
Оглавление
Введение
1.Энергия в экологических системах
1.1Принципы и концепции
2.Метаболизм и размер особей
2.1 Объяснения и примеры
Заключение
Литература
Введение
Термин «экология» (от греч. «Ойкос» – дом, жилище и «логос» – наука) был предложен более 100 лет назад выдающимся немецким естествоиспытателем Эрнстом Геккелем.
В буквальном смысле экология – это наука об условиях существования живых организмов, их взаимодействиях между собой и окружающей средой.
Экология – также междисциплинарное системное научное направление [27, 32]. Возникнув на почве биологии, оно включает в себя концепции, технологии математики, физики, химии. Но экология и гуманитарная наука, поскольку от поведения человека, его культуры во многом зависит судьба биосферы, а вместе с ней и человеческой цивилизации.
В зависимости от специфики решаемых экологических задач существуют ее разнообразные прикладные направления: инженерная, медицинская, химическая, космическая экология, агроэкология, экология человека и т.д.
Что является предметом исследования экологии? Экология изучает организацию и функционирование живых систем более сложных, чем организм, т. е. надорганизменных систем. Эти системы получили название экологических систем или экосистем.
Экосистема – это безразмерная устойчивая система живых и неживых компонентов, в которой совершается внешний и внутренний круговорот вещества и энергии [27]. В качестве примеров можно привести лесные экосистемы, почвы, гидросферу и т.д.
Самой крупной экосистемой, предельной по размерам и масштабам, является биосфера. Биосферой называют активную оболочку Земли, включающую все живые организмы Земли и находящуюся во взаимодействии с неживой средой (химической и физической) нашей планеты, с которой они составляют единое целое. Биосфера нашей планеты существует 3 млрд. лет, она растет и усложняется наперекор тенденциям холодной энтропийной смерти; она несет разумную жизнь и цивилизацию. Биосфера существовала задолго до появления человека и может обойтись без него. Напротив, существование человека невозможно без биосферы.
Все остальные экосистемы находятся внутри биосферы и являются ее подсистемами. Крупная региональная экосистема, характеризующаяся каким-либо основным типом растительности, называется биомом. Например, биом пустыни или влажного тропического леса. Гораздо меньшей системой является популяция, включающая группу особей одного вида, т. е. единого происхождения, занимающая определенный участок. Более сложной системой, чем популяция, является сообщество, которое включает все популяции, занимающие данную территорию. Таким образом, популяция, сообщество, биом, биосфера располагаются в иерархическом порядке от малых систем к крупным.
Одной из задач экологии является изучение превращения энергии внутри экологической системы. Усваивая солнечную энергию, зеленые растения создают потенциальную энергию, которая при потреблении пищи организмами превращается в другие формы. Превращения энергии в отличие от цикличного движения веществ идут в одном направлении, почему и говорят о потоке энергии.
1. Энергия в экологических системах
С точки зрения изучения потоков энергии важны два начала термодинамики. Первое начало гласит, что энергия не может создаваться заново и исчезать, а только переходит из одной формы в другую. Второе начало формулируется таким образом: процессы, связанные с превращениями энергии, могут протекать самопроизвольно лишь при условии, что энергия переходит из концентрированной формы в рассеянную. То, что согласно второму началу энергия при любых превращениях стремится перейти в тепло, равномерно распределенное между телами, дало основания говорить о «старении» Солнечной системы. Характерна ли эта тенденция к энергетическому выравниванию для всей Вселенной, пока не ясно, хотя в XIX веке широко обсуждался вопрос о «тепловой смерти Вселенной».
Общепринятая в физике формулировка второго начала гласит, что в закрытых системах энергия стремится распределиться равномерно, т. е. система стремится к состоянию максимальной энтропии. Отличительной же особенностью живых тел, экосистем и биосферы в целом является способность создавать и поддерживать высокую степень внутренней упорядоченности, т. е. состояния с низкой энтропией.
По определению Э. Шредингера, «жизнь – это упорядоченное и закономерное поведение материи, основанное не только на одной тенденции переходить от упорядоченности к неупорядоченности, но и частично на существовании упорядоченности, которая поддерживается все время... средство, при помощи которого организм поддерживает себя постоянно на достаточно высоком уровне упорядоченности (равно на достаточно низком уровне энтропии), в действительности состоит в непрерывном извлечении упорядоченности из окружающей его среды. В самом деле, у высших животных мы достаточно хорошо знаем тот вид упорядоченности, которым они питаются, а именно: крайне хорошо упорядоченное состояние материи в более или менее сложных органических соединениях служит им пищей. После использования животные возвращают эти вещества в деградированной форме, однако не вполне деградированной, так как их еще могут употреблять растения. Для растений мощным источником „отрицательной энтропии“, конечно, является солнечный свет» (Э. Шредингер. Что такое жизнь? С точки зрения физика. М., 1972, с. 71, 76).
Свойство живых систем извлекать упорядоченность из окружающей среды дало основания некоторым ученым, в частности Э. Бауэру, сделать вывод, что для этих систем второе начало не выполняется. Но второе начало имеет еще и другую, более общую формулировку, справедливую для открытых, в том числе живых, систем. Она гласит, что эффективность самопроизвольного превращения энергии всегда меньше 100 %. В соответствии со вторым началом поддержание жизни на Земле без притока солнечной энергии невозможно. «Все, что происходит в природе, означает увеличение энтропии в той части Вселенной, где это имеет место. Так и живой организм непрерывно увеличивает свою энтропию, или, иначе, производит положительную энтропию, и, таким образом, приближается к опасному состоянию – максимальной энтропии, – представляющему собой смерть. Он может избежать этого состояния, т. е. оставаться живым, только постоянно извлекая из окружающей среды отрицательную энтропию» (Там же, с. 76).
В экосистемах перенос энергии пищи от ее источника – растений через ряд организмов, происходящий путем поедания одних организмов другими, и называется пищевой цепью. При каждом очередном переносе большая часть (80–90 %) потенциальной энергии теряется, переходя в тепло.
Это ограничивает возможное число звеньев цепи до четырех-пяти. Зеленые растения занимают первый трофический уровень, травоядные – второй, хищники – третий и т. д. Переход к каждому следующему звену уменьшает доступную энергию примерно в 10 раз. Переходя к человеку, можно сказать, что если увеличивается относительное содержание мяса в рационе, то уменьшается число людей, которых можно прокормить.
Экологическая пирамида, представляющая собой трофическую структуру, основанием которой служит уровень продуцентов, а последующие уровни образуют ее этажи и вершину, может быть трех основных типов: «1) пирамида чисел, отражающая численность отдельных организмов; 2) пирамида биомассы, характеризующая общий сухой вес, калорийность или другую меру общего количества живого вещества; 3) пирамида энергии, показывающая величину потока энергии и (или) „продуктивность“ на последовательных трофических уровнях» (Ю. Одум. Основы... с. 105). Энергетическая пирамида всегда сужается кверху, поскольку энергия теряется на каждом последующем уровне.
Важнейшей характеристикой экосистемы является ее продуктивность, под которой понимается как рост организмов, так и создание органического вещества. Поглощается лишь около половины всей лучистой энергии (в основном в видимой части спектра), и самое большое около 5 % ее в самых благоприятных условиях превращается в продукт фотосинтеза. Значительная часть (не менее 20 %, а обычно около 50 %) этой потенциальной пищи (чистой продукции) человека и животных расходуется на дыхание растений. Содержание хлорофилла на 1 м2в разных сообществах примерно одинаково, т. е. в целых сообществах содержание зеленого пигмента распределено более равномерно, чем в отдельных растениях или их частях.
Соотношение между зелеными и желтыми пигментами можно использовать как показатель отношения гетеротрофного метаболизма к автотрофному. Когда в сообществе фотосинтез превышает дыхание, доминируют зеленые пигменты, а при усилении дыхания сообщества увеличивается содержание желтых пигментов.
Среди произведенной в процессе фотосинтеза продукции выделяют первичную продуктивность, которая определяется как скорость, с которой лучистая энергия усваивается организмами-продуцентами, главным образом зелеными растениями. Ее разделяют на валовую первичную продукцию, включая ту органику, которая была израсходована на дыхание, и чистую первичную продукцию – за вычетом использованной при дыхании растений. Чистая продуктивность сообщества – скорость накопления органического вещества, не потребленного гетеротрофами. Наконец, скорость накопления энергии на уровне консументов называют вторичной продуктивностью. В соответствии со вторым началом поток энергии с каждой ступенью уменьшается, так как при превращениях одной формы энергии в другую часть энергии теряется в виде тепла. «В более плодородных прибрежных водах первичная продукция приурочена к верхнему слою воды толщиной около 30 м, а в более чистых, но бедных водах открытого моря зона первичной продукции может простираться вниз на 100 м и ниже. Вот почему прибрежные воды кажутся темно-зелеными, а океанские – синими» (Там же, с. 70).
Часть энергии, идущая на дыхание, т. е. на поддержание структуры, велика в популяциях крупных организмов и в зрелых сообществах. Эффективность природных систем много ниже КПД электромоторов и других двигателей. В живых системах много «горючего» уходит на «ремонт», что не учитывается при расчете КПД двигателей. Любое повышение эффективности биологических систем оборачивается увеличением затрат на их поддержание. Экологическая система – это машина, из которой нельзя «выжать» больше, чем она способна дать. Всегда наступает предел, после которого выигрыш от роста эффективности сводится на нет ростом расходов и риском разрушения системы.
Человек не должен стремиться получать более одной трети валовой (или половины чистой) продукции, если он не готов поставлять энергию для замены тех «механизмов самообслуживания», которые развились в природе, чтобы обеспечить долговременное поддержание первичной продукции в биосфере. Прямое удаление человеком или домашними животными более 30–50 % годового прироста растительности может уменьшить способность экосистемы сопротивляться стрессу.
Один из пределов биосферы – валовая продукция фотосинтеза, и под него человеку придется подгонять свои нужды, пока не удастся доказать, что усвоение энергии путем фотосинтеза можно сильно повысить, не подвергая при этом опасности нарушить равновесие других, более важных ресурсов жизненного круговорота.
Урожай, получаемый человеком, составляет 1 % чистой или 0,5 % общей первичной продукции биосферы, если учитывать только потребление пищи человеком. Вместе с домашними животными это 6 % чистой продукции биосферы или 12 % чистой продукции суши.
Энергия, которую расходует человек, чтобы получить больший урожай, называется добавочной энергией. Она необходима для индустриализованного сельского хозяйства, так как этого требуют культуры, созданные специально для него. «Индустриализованное (использующее энергию горючих ископаемых) сельское хозяйство (как, например, практикуемое в Японии) может дать в 4 раза более высокий урожай с гектара, чем сельское хозяйство, в котором всю работу выполняют люди и домашние животные (как в Индии), но оно требует в 10 раз больших затрат разного рода ресурсов и энергии» (Там же, с. 526). Так называемые энергетические «субсидии» соответствуют закону убывающей отдачи А. Тюрго – Т. Мальтуса, формулируемому следующим образом: «Повышение удельного вложения энергии в агросистему не дает адекватного пропорционального увеличения ее продуктивности (урожайности)».
Замкнутость производственных циклов по энергетически-энтропийному параметру теоретически невозможна, поскольку течение энергетических процессов (в соответствии со вторым началом термодинамики) сопровождается деградацией энергии и повышением энтропии природной среды. Действие второго начала термодинамики выражается в том, что превращения энергии идут в одном направлении в отличие от цикличного движения веществ.
В формулировке Ю. Одума второе начало термодинамики справедливо по крайней мере для современного состояния системы «человек – природная среда», поскольку существование этой системы полностью зависит от притока солнечной энергии. Мы являемся свидетелями того, что повышение уровня организации и разнообразия культурной системы уменьшает ее энтропию, но увеличивает энтропию окружающей природной среды, вызывая ее деградацию. В какой степени можно элиминировать эти следствия второго начала? Существуют два пути. Первый заключается в уменьшении потерь используемой человеком энергии при ее различных превращениях. Этот путь эффективен в той мере, в которой не приводит к понижению стабильности систем, через которые идет поток энергии (как известно, в экологических системах увеличение числа трофических уровней способствует повышению их устойчивости, но в то же время росту потерь энергии, проходящей через систему). Второй путь заключается в переходе от повышения упорядоченности культурной системы к повышению упорядоченности всей биосферы. Общество в этом случае повышает организованность природной среды за счет понижения организованности той части природы, которая находится за пределами биосферы Земли.
экологический система энергия метаболизм
1.1 Концепция экосистемы
Живые организмы и их абиотическое окружение неразделимо связаны друг с другом и находятся в постоянном взаимодействии. Любая биологическая система, включающая все совместно функционирующие организмы (биотическое сообщество) на данном участке и взаимодействующая с физической средой таким образом, что поток энергии создает четко определенную трофическую структуру, видовое разнообразие и круговорот веществ между живой в неживой частями, представляет собой экологическую систему или экосистему.
Долговременное функционирование экосистемы обеспечивают три основных компонента - сообщество, поток энергии и круговорот веществ.
Поток энергии направлен в одну сторону; часть поступающей солнечной энергии преобразуется сообществом и переходит на качественно более высокую ступень, трансформируясь в органическое вещество, представляющее собой более концентрированную форму энергии, чем солнечный свет; но большая часть энергии деградирует, проходит через систему и покидает ее в виде низкокачественной тепловой энергии ("тепловой сток"). В конечном итоге эта судьба ожидает всю энергию, поступающую в биосферу. Энергия может накапливаться в экосистеме, затем снова высвобождаться или экспортироваться в другую систему, но ее нельзя использовать вторично.
В отличие от энергии элементы питания, в том числе биогенные элементы, необходимые для жизни (углерод, водород, кислород, азот, фосфор и др.), не только могут, но и должны использоваться многократно.
Все экосистемы, даже самая крупная - биосфера, являются открытыми системами: они должны получать и отдавать энергию. Разумеется, экосистемы, входящие в биосферу, также в разной степени открыты для потоков веществ, для иммиграции и эмиграции организмов. Поэтому концепция экосистемы должна учитывать существование связанных между собой и необходимых для функционирования и самоподдержания экосистемы "среды на входе" и "среды на выходе": в концептуально законченную экосистему входит среда на входе, среда на выходе и система , т.е.:
Экосистема=IE+S+OE,
где IE - среда на входе; S - система; OE - среда на выходе.
Данная схема решает проблему, связанную с
проведением границ рассматриваемой единицы, поскольку в этом случае не имеет
значения, как мы вычленяем исследуемую часть экосистемы. Часто удобными
оказываются естественные границы, например берег озера или опушка леса; или
административные, например границы города; но эти границы могут быть и
условными, если их точно определить геометрически. Конечно, экосистема не
ограничена "ящиком" в центре схемы, поскольку если бы этот
"ящик" был герметичным, то его живое содержимое (озеро или город) не
вынесло бы такого заключения. Функционирующая реальная экосистема должна иметь
вход и в большинстве случаев пути оттока переработанной энергии и веществ.
2. Метаболизм и размер особей
Под метаболизмом понимают постоянно происходящий в клетках живых организмов обмен веществ и энергии. Одни соединения, выполнив свою функцию, становятся ненужными, в других возникает насущная потребность. В различных процессах метаболизма из простых веществ при участии ферментов синтезируются высокомолекулярные соединения, в свою очередь сложные молекулы расщепляются на более простые.
При неизменном энергетическом потоке в пищевой цепи более мелкие организмы имеют более высокую интенсивность обмена, более высокий удельный метаболизм (метаболизм в пересчете на 1 кг массы), чем крупные организмы. При этом мелкие организмы создают относительно меньшую биомассу, чем крупные. Так, биомасса бактерий, имеющихся в данный момент в экосистеме, гораздо ниже биомассы млекопитающих. Эта закономерность получила название правила Одума. Это правило заслуживает особого внимания, поскольку из-за антропогенного нарушения природы происходит измельчание организмов, которое неминуемо должно привести к общему снижению продуктивности и к разладу в экосистемах.
При измельчании особей выход биомассы с единицы площади в силу более плотного заселения пространства увеличивается. Слоны не дадут такой биомассы и продукции с единицы площади, которую способна дать саранча. Это – закон удельной продуктивности. Так, мелкие предприятия и фермы в сумме производят больший объем хозяйственной продукции, чем крупные, тем более крупнейшие.
Исчезновение видов, представленных крупными особями, меняет структуру экосистем. При этом организмы одной трофической группы замещают друг друга. Так, копытных в степи и саванне сменяют грызуны, а в ряде случаев – растительноядные насекомые. Это – принцип экологического дублирования.
В результате потери энергии при переносе ее по трофической цепи и таких факторов, как зависимость метаболизма от размеров особи, каждая экосистема приобретает определенную трофическую структуру. Ее можно представить в виде экологических пирамид. Если принять, что в вещество тела животного переходит в среднем 10% энергии съеденной пищи, то за счет 1 т растительной массы может образоваться 100 кг массы тела травоядного животного, а за счет последнего – 10 кг массы тела хищников.
2.1 Обьяснения и примеры
У мелких растений и животных — водорослей, бактерии, простейших — удельный метаболизма (на 1 г биомассы) значительно выше, чем у крупных, например деревьев и позвоночных. Это относится и к фотосинтезу, и дыханию. Во многих случаях самыми важными для метаболизма сообщества оказывается не малочисленные крупные, выделяющиеся своим размером организмы, а многочисленные мельчайшие организмы, часто не различаемые невооруженным глазом. Например, метаболизм микроскопических водорослей (фитопланктона), которых в озере в каждый данный момент времени наберется не более нескольких килограммов на гектар, может не отличаться от метаболизма намного большей биомассы деревьев в лесу или травы на лугу. Точно так же несколько килограммов мелких ракообразных (зоопланктон), «пасущихся» на водорослях, могут иметь общее дыхание, равное дыханию многих килограммов пасущейся на лугу коровы.
Интенсивность обмена у отдельных организмов или их ассоциаций часто оценивается по скорости потребления кислорода (или в случае фотосинтеза — но скорости его продукции). У животных наблюдается общая тенденция к увеличению интенсивности обмена в расчете на организм пропорционально степени 2/з роста объема (или массы); иначе говоря, интенсивность метаболизма на грамм биомассы растет с уменьшением длины (Zeuthen, 1953; Bertalanffy, 1957; Klеiber, 1961).
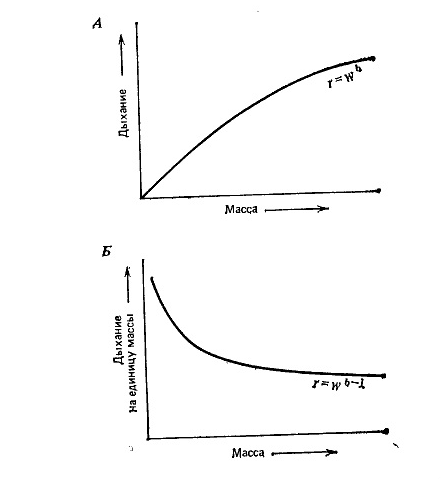
Рис. 3-17. Связь между дыханием и массой тела у одной особи (А) и дыханием на единицу массы и массой тела (Б). Показатель степени в уравнениях обычно лежит в пределах 0,7—0,8. (С изменениями по Agren, Axelsson, 1980.)
Видимо, подобное соотношение наблюдается и у растений, хотя различия и строении растений и животных затрудняют прямое сравнение объемов и линейных размеров. Связь между массой тела и интенсивностью дыхания на особь и на единицу массы показана на рис. 3,17» Для нас представляет интерес вторая кривая (рис. 3,17, Б), поскольку она показывает рост удельной (по массе) интенсивности метаболизма с уменьшением размеров особи. Различные теории, объясняющие эту тенденцию, особое внимание обращают на роль процессов диффузии; действительно, у крупных организмов площадь поверхности, через которую может идти диффузия, на грамм массы меньше, чем у мелких животных. Однако общепринятого объяснения связи между размером и метаболизмом пока нет. Конечно, сравнения следует проводить при одинаковой температуре, так как интенсивность обмена при высокой температура обычно выше (кроме случаев температурной адаптации; см. с 261 и 202),
При сравнении организмов, имеющих размеры одного порядка, линейные зависимости, показанные на рис. 3.17, не всегда оказываются справедливыми. Это вполне естественно, потому что на интенсивность метаболизма влияют и многие другие факторы. Например, у теплокровных животных интенсивность дыхания выше, чем у холоднокровных такого же размера. По это различие относительно мало но сравнению с различием между позвоночными и бактериями. Итак, при одинаковом поступлении энергии с пищей величины урожая па корню холоднокровной растительноядной рыбы в пруду и урожая теплокровных травоядных млекопитающих на суше могут быть одного порядка. Но, как ужо указывалось в гл. 2, в воздухе кислорода больше, чем в воде, где он может служить лимитирующим фактором. Б общем у водных животных, по-видимому, активность дыхания на единицу массы меньше, чем у наземных животных такого же размера. Такая адаптация вполне может повлиять на трофическую структуру
При анализе связи размеров с метаболизмом у растений часто нелегко решить, что же, собственно говоря, считать «особью». Так, большое дерево можно рассматривать как одну особь, по при изучении связи размеров с площадью поверхности «функциональными особями» можно считать листья (вспомним понятие «индекс листовой поверхности»). Изучая разные виды крупных морских многоклеточных водорослей, мы обнаруживаем, что виды с тонкими, или узкими, «ветвями» (т. е. с высоким соотношением поверхность/объем) характеризуются более высоким уровнем продукции пищи на 1 г биомассы, более интенсивным дыханием и поглощением радиоактивного фосфора из воды, чем виды с толстыми «ветвями» (Е. Odum, Kuentzler, Blunt, 1958). В этом случае «функциональными особями» являются «ветви* или даже отдельные клетки, а не все растение, которое может быть образовано множеством «ветвей», прикрепленных к субстрату одним ризоидом.
В онтогенезе любого вида также может наблюдаться обратная зависимость между размерами тела и интенсивностью метаболизма. Так, в яйцах интенсивность метаболизма на 1 г массы обычно выше, чем у взрослых особей. По данным Хантера и Вернберга (Hunter, Vernberg, 1955), метаболизм на 1 г массы у взрослых трематод (паразитических червей) в 10 раз ниже, чем у их мелких личинок церкарий.
Заключение
И так переходим к заключению. Вывод можно сделать такой. Что размер урожая биомассы на корню (выраженный в общем сухом весе или обшей калорийности всех организмов, имеющихся в данный момент), который может поддерживаться постоянным током энергии через пищевую цепь, в значительной мере зависит от размера особей. Чем меньше организм, тем выше его удельный метаболизм (на 1 г или на 1 кал биомассы). Следовательно, чем меньше организм, тем меньше биомасса, которая может поддерживаться на данном трофическом уровне экосистемы, и, наоборот, чем крупнее организм, тем выше биомасса на корню. Так, «урожай» бактерий, имеющихся в данный момент, будет гораздо ниже «урожая» рыбы или млекопитающих, хотя эти группы использовали одинаковое количество энергии.
Во избежание недоразумений подчеркиваем, что с увеличением размера снижается не общий метаболизм особи, а удельный, т.е. интенсивность метаболизма не единицу веса. Взрослому человеку требуется пищи больше, чем маленькому ребенку, но на килограмм своего веса взрослый потребляет меньше пищи.
Литература
Ю. Одум, 1975 г. «Основы экологии».
Сайт: http://www.xumuk.ru/
| Концепции современного естествознания | |
|
СМОЛЕНСКИЙ ИНСТИТУТ БИЗНЕСА И ПРЕДПРИНИМАТЕЛЬСТВА Сычев М.М. КОНЦЕПЦИИ СОВРЕМЕННОГО ЕСТЕСТВОЗНАНИЯ СОДЕРЖАНИЕ Введение. Две культуры как отражение ... Продукты метаболизма (обмена) с высоким содержание энтропии организм выделяет в окружающую среду. Продуктивность биосферы представляет собой биомассу, производимую различными экосистемами, составляющими биосферу. |
Раздел: Рефераты по биологии Тип: учебное пособие |
| Физиология растений | |
|
Куниченко Наталья Александровна, кандидат сельскохозяйственных наук, доцент, заведующая кафедрой защиты растений и экологии Приднестровского ... Дыхание является ключевым процессом метаболизма любого организма по двум причинам: при дыхании происходит освобождение химической энергии органических веществ, используемых в ... Оценка эффективность накопления растением биомассы определяется рядом показателей: суточным приростом биомассы растения, коэффициентом эффективности роста, соотношением фотосинтеза ... |
Раздел: Рефераты по биологии Тип: учебное пособие |
| Атлантический океан: биогеоценоз и экологические проблемы | |
|
Курсовая работа по физической географии Сдавалась на географическом факультете Кубанского государственного университета. 1. Введение. Атлантический ... Совокупность всех живых организмов биосферы, существующих в данный момент, численно выраженных в элементарном химическом составе в весе и энергии, В. И. Вернадский назвал живым ... Одинаковая продукция сельди при различной интенсивности использования ее запасов в какой-то мере подтверждает наши предположения о том, что старшие возрастные группы рыб не ... |
Раздел: Рефераты по географии Тип: дипломная работа |
| Билеты по биологии для 10-11 классов | |
|
Билеты по биологии БИЛЕТ №1 ВОПРОС 1. Уровни организации живой материи Молекулярный. Любая живая система, как бы сложно она ни была организована ... Экология занимается изучением отдельных особей, популяций (состоящих из особей одного вида), сообществ (состоящих из популяций), и экосистем (включающих сообщества и окружающую их ... Биосфера. Совокупность всех биогеоценозов (экосистем) Земли представляет собой большую экологическую систему - биосферу. |
Раздел: Биология и химия Тип: шпаргалка |
| Ответы 11 класс | |
|
БИЛЕТ №1 1. Биосфер, границы биосферы, глобальные проблемы биосферы. Биосфера - это самая большая экосистема. Это оболочка планеты, заселённая живыми ... В наземных экосистемах с повышением трофического уровня она убывает, потому что энергия теряется при переходе с одного трофического уровня на другой. В наземных экосистемах с повышением трофического уровня запас биомассы уменьшается, а в морских - увеличивается. |
Раздел: Рефераты по экологии Тип: реферат |