Реферат: Изучение миксомицетов среднего Урала, выращенных методом влажных камер
Уральский государственный университет
им. А.М. Горького
Кафедра ботаники
РЕФЕРАТ
Изучение миксомицетов среднего Урала, выращенных методом влажных камер
Исполнитель Руководители
Студент гр.Б-101 проф. Мухин В.А.
Плотников Б.С. Ушакова Н.В.
Фефелов К.А.
Екатеринбург 2001
СОДЕРЖАНИЕ
I.БИБЛИОГРАФИЧЕСКАЯ СПРАВКА
1. Отдел слизевики
1.1. Класс протостелиевые
1.2. Класс плазмодиофоровые
1.3. Класс клеточные слизевики или акразиевые
2. Класс собственно слизевики
2.1. Жизненный цикл
2.2. Классификация
2.3. Сбор в природе
2.4. Выращивание культур. Метод влажных камер
2.5. Экология Myxomycetes
2.6. Географическое распределение
2.7. Структурные характеристики Myxomycetes
3. Ученые, работавшие в области миксомикологии
II. РЕЗУЛЬТАТЫ ПРАКТИЧЕСКИХ ИССЛЕДОВАНИЙ
Постановка эксперимента
Описание образцов
Методика эксперимента
Результаты
Определение видов
Характерные особенности обнаруженных видов
Выводы
ПРИЛОЖЕНИЕ 1
ПРИЛОЖЕНИЕ 2
СПИСОК ЛИТЕРАТУРЫ
Библиографическая справка
Отдел слизевики Myxomycota.
Отдел объединяет низших эукариотических организмов, довольно разнообразных по строению и образу жизни. Среди них есть как свободноживущие в почве, на навозе, на разлагающихся растительных остатках и тому подобных субстратах, так и паразиты водорослей, водных грибов и высших водных и наземных растений.[1]
Некоторые из них имеют микроскопически малые размеры и представлены одноядерной или многоядерной амебоидной клеткой, у других вегетативное тело в виде крупного многоядерного цитоплазменного образования, так называемого плазмодия, достигающего в отдельных случаях даже нескольких десятков сантиметров.
В вегетативной стадии большинство слизевиков, представленных амебоидными клетками, размножается делением этих клеток.
Отдел включает следующие классы, различающиеся по степени организации и особенностям циклов развития своих представителей: ПРОТОСТЕЛИЕВЫЕ (Protosteliomycetes), cобственно слизевики (Myxomycetes), плазмодиофоровые (Plasmodiophoromycetes), акразиевые, или клеточные слизевики (Acrasiomycetes).
1.1 Kласс ПРОТОСТЕЛИЕВЫЕ - Protosteliomycetes
Большинство представителей класса обнаружено и описано только в середине нашего столетия, хотя и представляет собой широко распространенные организмы. Они могут быть выделены из отмершей древесины, коры деревьев, из почвы, особенно богатой гумусом, из навоза, из покровов отмерших, но не отделившихся плодов (коробочек, стручков, ягод и т. п.).
В настоящее время известно около десятка родов и немногим более 65 видов этого класса. Большинство представителей относительно просты по морфологии и характеризуются микроскопически малыми размерами вегетативных тел—амебоидов.
Из амеб (или частей плазмодия у некоторых протостелиевых) образуются плодовые тела—спорокарпы непосредственно на субстрате, а у других — на рожкообразных выростах. Они представляют собой сидящую на тонкой полой ножке головку с одной, двумя или большим числом спор. Зрелая головка отделяется от ножки, и в подходящих условиях споры освобождаются и прорастают четырьмя—восемью одноядерными подвижными клетками — зооспорами, превращающимися в амебопды, или непосредственно амебоидами, с которых начинается новый цикл. У видов, имеющих плазмодий, последний развивается из разрастающихся амебоидных клеток.
По-видимому, протостелиевые в своем происхождении связаны со свободноживущими жгутиковыми (Flagellatae), на что указывает существование жгутиковой стадии у некоторых представителей. У бесцветных и окрашенных жгутиковых также наблюдается образование плазмодиев и оно, безусловно, облегчало выход на сушу древним жгутиковым. Развитие спороношений было дальнейшим шагом приспособления к наземной жизни. Наиболее вероятно, что миксомицеты, как и Flagellata произошли от общих жгутиковых проформ.
1.2.КЛАСС ПЛАЗМОДИОФОРОВЫЕ — PLASMODIOPHOROMYCETES.
Представители класса — внутриклеточные паразиты. В отличие от свободноживущих слизевиков они не образуют никаких специальных спороношений, а споры развиваются из плазмодия прямо внутри клетки хозяина.
Наиболее известный представитель — возбудитель килы капусты и других крестоцветных—плазмодиофора капустная {Plasmodiophora brassicae). Пораженные корни растения имеют ненормальный, уродливый вид, который придают им объемистые опухоли . Заболевание очень вредоносно, так как при заражении капустной рассады кочаны могут совсем не образоваться, а при заражении взрослых растений они недоразвиваются. На срезах корней больных растений в клетках коровой паренхимы видны или небольшие плазменные комочки — миксамебы, или плазмодии, часто заполняющие весь объем клетки. На поздних стадиях развития болезни плазмодий распадается на массу мелких круглых спор. Зараженные клетки паренхимы корня по размеру больше здоровых и расположены группами. Причина этого в том, что паразит, находясь в клетке, вызывает увеличение размеров клеток (гипертрофию) и усиленное их деление (гиперплазию), при котором паразит оказывается и в дочерних клетках. За счет таких реакций клеток на заражение и развиваются опухолевые разрастания корней. Более тонкий механизм образования опухоли заключается в нарушении синтеза фенольных и индольных соединений в растении под влиянием паразита.
При сгнивании корней споры попадают в почву. Их распространению в почве способствуют животные (дождевые черви, насекомые), токи воды (дождь, полив), деятельность человека, например перенос земли со спорами на орудиях обработки почвы, и т. п. В почве споры могут годами сохраняться, не теряя способности к прорастанию. При подходящих условиях влажности и температуры и при стимулирующем действии корневых выделений растений-хозяев споры прорастают.
Образующиеся из них зооспоры, или миксамебы, заражают новые растения главным образом через корневые волоски. В клетке корневого волоска происходит слияние миксамеб (плазмогамия), ведущее к формированию первичного плазмодия с гаплоидными ядрами, число которых несколько увеличивается за счет митотических делений. Из такого плазмодия развиваются гаметангии или зооспорангии и в них—зооспоры (или гаметы), выходящие из корневого волоска в почву. Они попарно сливаются (без слияния ядер). Образовавшаяся двухъядерная клетка заражает корень капусты и дает начало новым, более крупным вторичным плазмодиям. В них многочисленные ядра, образующиеся в результате митотических делений, попарно сливаются, затем следует редукционное деление диплоидных ядер и плазмодий распадается на споры. Таким образом, часть цикла развития паразита осуществляется в корневых волосках, часть—в почве, а еще одна часть—в клетках корня. Необходимо отметить, что зооспоры плазмодиофоры, как и зооспоры истинных слизевиков, имеют два гладких жгутика неравной длины. Впервые капустную килу тщательно изучил М. С. Воронин (1878). Он установил причину заболевания, описал в главных чертах цикл развития паразита и предложил меры борьбы с ним, а также указал на его способность заражать другие растения из семейства крестоцветных.
Другой представитель этого класса, имеющий важное практическое значение,—возбудитель порошистой парши картофеля - спонгоспора (Spongospora solani). Многоядерные плазмодии спонгоспоры развиваются в периферических клетках клубня и образуют затем губчатые комочки плотно спаянных спор. Пораженный клубень покрыт язвочками с коричневым порошистым содержимым, состоящим из скоплений этих комочков. Иногда развивается корневая форма в виде небольших белых желвачков, темнеющих и затем распадающихся. Кроме картофеля поражаются томаты и другие виды пасленовых.
Филогенетические связи представителей этого класса еще недостаточно ясны. Некоторые ученые сближают их с простейшими животными; другие, основываясь на чертах сходства в их строении и развитии с примитивными грибами—внутриклеточными паразитами, относят их к грибам; третьи, исходя из особенностей жгутиковых стадий, сближают их со слизевиками, считая этот класс уклонившейся ветвью развития слизевиков, изменения в которой, в частности утрата соответствующих спороношений, произошли в результате паразитического образа жизни. При этом общие черты с некоторыми примитивными грибами следует рассматривать как конвергенцию под влиянием сходных условий существования.
1.3. КЛАСС КЛЕТОЧНЫЕ СЛИЗЕВИКИ, ИЛИ AKPA3ИЕВЫЕ—ACRASIOMYCETES (Dictyostelids)
Представители класса, небольшого по числу видов (несколько более двух десятков), обитают на гниющих растительных остатках, преимущественно на навозе, в почве. Вегетативная стадия представлена свободноживущими одноядерными амебами, питающимися фаготрофно и размножающимися делением. По исчерпании запасов пищи амебы сползаются (стадия агрегации), но полностью не сливаются (хотя в центре скопления возможны плазменные связи между ними), образуя псевдоплазмодий. У многих представителей псевдоплазмодий перемещается (стадия миграции), постепенно формируя спороношение в виде головки из спор, сидящей на ножке (стадия кульминации). Из спор снова выходят амебы.
Наиболее известный представитель—диктиостелиум (Dictyostеlium discoideum). Его нежная прозрачная шаровидная головка из спор, одетых целлюлозной оболочкой, диаметром 0,2—0,3 мм сидит на прямой беловатой или желтоватой ножке длиной 3—8 мм. Он хорошо развивается в культуре (на агаризованном сенном отваре с кишечной палочкой в качестве источника пищи) и проходит весь цикл развития за 3—4 дня. Это обстоятельство сделало его одним из излюбленных объектов экспериментальных исследований (выяснение механизмов различных стадий онтогенеза, дифференциации, таксисов, возникновения гетерокариоза при агрегации миксамеб и др.).
Можно предположить филогенетическую связь акразиевых с протостелиевыми или со свободноживущими почвенными амебами.
КЛАСС СОБСТВЕННО СЛИЗЕВИКИ – MYXOMYCETES
ПОДКЛАСС MYXOGASTEROMYCETIDAE
Название myxomycetes получили от Греческих слов myxa (слизь) и myketes (грибы), и сначала использовалось в 1833, чтобы описать группу. Однако, некоторые биологи чувствовали, что слизевики более животно-подобны, называя их Mycetezoa, от Греческого mykes (гриба) и zoon (животное).[2]
Myxomycetes, обычно называемые слизевиками, являются одними из наиболее экстраординарных организмов , т.к. они проявляют свойства и, грибов и животных.
ЖИЗНЕННЫЙ ЦИКЛ
Жизненный цикл начинается со споры
(A), образованной в спорофоре. При соответствующей влажности и температурных
условиях, 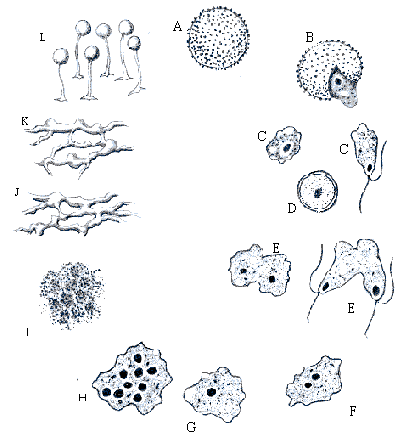 спора прорастет и произведет 1-4 протопласта. (B)
Протопласты (C), выходящие через трещины или поры в стенках споры, могут быть
флагеллятными клетками или бесформенными миксамебами. Если влажность
достаточная формируются флагеллятные клетки. В сухой окружающей среде -
миксамебы. Обычно флагеллятные клетки появляются прямо из споры и могут
превращаться в миксамебы. В случае неблагоприятных условий роста, они могут
стать микрокистами (D), чтобы выжить в течение более длинных периодов времени.
Слияние 2 совместимых клеток (E), или флагеллятных или миксамеб (плазмагамия и кариогамия), заканчивается образованием
диплоидной зиготы (F и G). После периода питания и роста, зигота развивается в
одноклеточную, многоядерную структуру (H), плазмодий. При неблагоприятных
условиях может образоваться другая стойкая структура, склероций (I). При
благоприятных условиях склероций может преобразовываться в плазмодий (J).
Зрелый плазмодий (K) производит один или большее количество споросодержащих
структур, называемых плодовыми телами(L).
спора прорастет и произведет 1-4 протопласта. (B)
Протопласты (C), выходящие через трещины или поры в стенках споры, могут быть
флагеллятными клетками или бесформенными миксамебами. Если влажность
достаточная формируются флагеллятные клетки. В сухой окружающей среде -
миксамебы. Обычно флагеллятные клетки появляются прямо из споры и могут
превращаться в миксамебы. В случае неблагоприятных условий роста, они могут
стать микрокистами (D), чтобы выжить в течение более длинных периодов времени.
Слияние 2 совместимых клеток (E), или флагеллятных или миксамеб (плазмагамия и кариогамия), заканчивается образованием
диплоидной зиготы (F и G). После периода питания и роста, зигота развивается в
одноклеточную, многоядерную структуру (H), плазмодий. При неблагоприятных
условиях может образоваться другая стойкая структура, склероций (I). При
благоприятных условиях склероций может преобразовываться в плазмодий (J).
Зрелый плазмодий (K) производит один или большее количество споросодержащих
структур, называемых плодовыми телами(L).
Все структуры, кроме миксамеб и флагеллятных клеток, диплоидны. Редукционное деление происходит при их образовании .
Некоторый миксомицеты развиваются без полового размножения, их жизненный цикл является полностью диплоидным. Некоторая виды могут использовать оба типа жизненных циклов или переходить от одного типа к другому.
Классификация
Отдел Myxomycota
Класс Myxomycetes
Подкласс Myxogasteromycetidae[3](рис. См. Приложение I)
|
ПОРЯДОК |
СЕМЕЙСТВО |
РОДЫ |
|
| Echinsteliales | Echinosteliaceae | Echinostelium | |
| Clastodermataceae |
Barbeyella Clastoderma |
||
| Liceales | Liceaceae | Licea | |
| Reticulariaceae |
Dictydiaethalium Lycogala Reticularia Tubifera |
||
| Cribrariaceae |
Cribraria Lindbladia |
||
| Trichiales | Dianemaceae |
Calomyxa Dianema Listerella Minakatella |
|
| Trichiaceae |
Arcyria Arcyodes Calonema Cornuvia Hemitrichia Metatrichia Oligonema Perichaena Prototrichia Trichia |
||
| Stemonitales | Stemonitaceae |
Amurochaete Brefeldia Colloderma Comatricha Diachea Diacheopsis Elaeomyxa Enerthenema Lamproderma Leptoderma Macbrideola Paradiacheopsis Stemonitis |
|
| Physarales | Physaraceae |
Badhamia Badhamiopsis Cienkowskia Craterium Erionema Fuligo Leocarpus Physarum Protophysarum |
|
| Didymiaceae |
Diderma Didymium Lepidoderma Mucilago |
||
2.3. Сбор в природе
В основном слизевики встречаются в природе в стадии хрупкого плодового тела. Чтобы собирать плодоносящие тела необходимо удалить часть подложки, на которой находится спорофор карманным ножом. Чтобы не повредить экземпляр надо использовать пластиковые или картонные контейнеры. Спичечные коробки идеальны для этой цели. Маленькие растительные частицы (листья, ветви папоротника, или мхи) могут быть помещены в контейнер на подушечке для сохранения в процессе транспортировки. Если сбор - плазмодии, в контейнер необходимо поместить увлажненное бумажное полотенце, или фильтровальную бумагу, чтобы развитие слизевика продолжалось. Каждый контейнер должен быть помечен, чтобы было известно, где и когда был проведен сбор.[2]
В лаборатории позвольте зрелым экземплярам высохнуть, удаляя влагу, конденсирующуюся на крышке контейнера. После высыхания, экземпляр можно вклеить в помеченную коробку для постоянного хранения. Ярлык должен содержать информацию о видовой принадлежности, подложке на который произошло плодоношение, местоположение, дату и имя собравшего. Можно указать порядковый номер и имя определившего.
Чтобы предотвратить повреждение насекомыми, коллекции могут быть помещены в домашний морозильник на нескольких дней.
2.4. Выращивание культур. Метод влажных камер.
Плодовые тела миксомицетов могут быть собраны в течение лета и начала осени в умеренных районах мира. Однако, наблюдение и сбор коллекций миксомицетов не ограничены этими сезонами и могут иметь место и в другие сезоны, при использовании простой техники выращивания культуры. Этот метод культуры был развит в 1933, для наблюдения за ростом водоросли Protococcus на коре дерева. Эта техника начиная с того времени стала использовться и в изучении миксомицетов.
2.4.1.Сбор коры
Чтобы приготовить влажные камеры, нужно сначала собрать маленькие (1-2 см) кусочки наружной коры с живого дерева с использованием ножа или отвертки. (Другие материалы типа листьев, прутов и экскрементов травоядных животных могут также использоваться и будут обсуждены позже.) При удалении коры, надо постараться не повредить дерево. Деревья типа дуба (Quercus), ясенья (Fraxinus), вяза (Ulmus), клена (Acer) и гикори (Carya) с грубой корой производительнее чем деревья с гладкой корой. Главным образом, кора хвои не очень производительная; исключение - можжевельник (Juniperus), хотя кора сосны используется очень часто. После сбора кусочки коры должны быть помещены в помеченные мешки. Вообще, пластиковые мешки хороши для краткосрочного хранения. Для хранения более длительное время, чем несколько дней, должны использоваться бумажные мешки.
2.4.2.Подготовка Влажных камер
Любой контейнер, который может быть закрыт крышкой, можно использовать, как влажную камеру. Одноразовая пластмассовая чашка Петри – подходящй контейнер . Она может быть выброшена после использования. Спорофоры миксомицетов могут развиваться на крышке или стенке влажной камеры. Когда используется пластмассовая чашка Петри, часть пластмассы с плодоносящим телом можно просто вырезать из посуды. Тем не менее неплохи и стеклянные чашки Петри.
Кусок фильтровальной бумаги или бумажного полотенца необходимо обрезать по размеру камеры и поместить в основание . Далее необходимо поместить образцы (кора, листья, или пруты) поверх фильтровальной бумаги так, чтобы они закрыли основание, но не накладывались друг на друга. Кора должна быть помещена внешней поверхностью вверх. Необходимо добавить достаточное количество дистиллированной воды чтобы образец был закрыт. Затем, закройте контейнер, и отложите его на сутки, чтобы вода впиталась.
Контейнер должен быть помечен типом образца и датой. Восковой карандаш или маркер идеальны для этой цели. На следующий день, вода должна быть слита и культуры должны остаться в рассеяном свете при комнатной температуре.
2.4.3.Просмотр Культур
Культуры должны быть проверены после нескольких дней и затем просматриваться регулярно каждые несколько дней по крайней мере две или три недели. Культуры могут сохраняться в течение нескольких месяцев, так как некоторый миксомицеты не развиваются в течение нескольких недель или даже дольше. Однако, некоторые миксомицеты с очень маленькими спорофорами, включая вид Echinostelium, иногда появляются в пределах от 24 до 48 часов.
Добавлять воду к культурам не надо. Это может привести к неправильному развитию спорофоров, а также к развитию плесени (различнах грибов), которые способны подавить развитие миксомицетов, даже уничтожить их.
Культуры должны быть исследованы тщательно, чтобы обнаружить все плодоносящие тела. Несколько виды могут развиваться в единственной культуре. Культуры можно проверять, используя лупу с увеличениеи Х16-54 или, если доступно, бинокулярная лупа. При проверке культур можно не снимать крышку с контейнера, но часто это необходимо сделать, так как она создает блики.
Если в чашки все-таки была добавлена вода, то найденные спорофоры необходимо удалять из культуры и фиксировать, иначе спорофор может быть уничтожен грибами. В сухих камерах такой опасности нет.
2.5.Экология Myxomycetes
Хотя развитие, генетика, географическое распределение, физиология и ультраструктура миксомицетов - обычные предметы исследования, относительно немного известно об их экологии. Большинство исследований экологии слизевиков было основано на наблюдениях репродуктивной стадии в жизненном цикле. Эта стадия обычно наблюдается в природе. Микроскопические амебофлагеллятные клетки (одноклеточная пищевая стадия) легко наблюдаются в лаборатории, но практически невозможно наблюдать их в полевых условиях. Плазмодий (многоядерная пищевая стадия), хотя макроскопический - обычно не готов к споруляции.
Миксомицеты - типичные обитатели наземных экосистем. Для развития и образования спорофоров они используют разнообразные субстраты: подстилка, разлагающаяся древесина, кора деревьев, помет растительноядных животных, а также любую другую органику растительного происхождения. Считается, что большинство видов не приурочено к определенному типу субстрата. Тем не менее некоторые виды и даже более крупные таксоны достаточно узко специализированы, как например р. Cribraria специализированный к древесине хвойных, а также р. Didymium, большинство представителей которого предпочитают развиваться на подстилке. Однако до сих пор нет данных о приуроченности отдельного вида миксомицета к определенной породе дерева или роду в целом. Дроздович (Drozdowicz, 1977), изучал субстратное распределение миксомицетов на различных древесных породах и отметил, что различия касаются только того к какому классу относится дерево — голосеменным или покрытосеменным.[4]
Как отмечалось многими авторами (Новожилов, 1993; Мир растений, 1991; Stephenson, 1994), наибольшее количество видов ассоциировано с древесиной. На сегодняшний день известно около тысячи видов миксомицетов (Ainthwors, Bisby, 1995), из них около 70% видов в той или иной степени приурочено к древесине. Однако, эта цифра приведена только для бореальных лесов. В растительных сообществах, где отсутствует или плохо развита подстилка, а также другие типы субстратов эта цифра может доходить до 90 и более процентов.
2.5.1. Трофические Стадии
Обе питающиеся стадии миксомицетов обычно существуют в микросредах обитания, где популяции бактерий высоки. Бактерии, вероятно, главный источник продовольствия для миксомицетных амебофлагеллятных клеток. Escherichia coli и Enterobactor aerogenes использовались как пищевой ресурс для лабораторных исследований. Амебофлагеллятные клетки, как известно, питаются различными спорами грибов (Гильберт 1928). Разумно предположить, что, так как эти клетки можно вырастить на химически определенных питательных средах в лаборатории, прямое поглощение растворенных органических питательных веществ происходит и в природе.
Плазмодии, как известно, питаются разнообразными бактериями (Cohen 1941), но не все бактерии подходят для развития плазмодия, и некоторые могут даже прекращать рост. Остатки бактериальных клеток в пищеварительных вакуолях могут быть вытеснены в стебелек при споруляции (Blackwell 1974). Используя электронную микроскопию были обнаружены бактериальные споры и другие частицы в стебельках семи видов миксомицетов.[2]
В природе плазмодии поглощают различные типы пищи, включая водорослевые клетки, дрожжи и споры, вегетативные гифы и (в некоторых случаях) плодоносящие тела грибов (Lister 1888, Говард и Curie 1932). Плазмодии также питаются плазмодиями других видов (McManus 1962). Кроме того, плазмодий способен абсорбировать питательные вещества непосредственно из окружающей среды, и некоторые виды могут показывать внеклеточное переваривание растительного и микробного материала.
Плазмодии хорошо приспособлены, чтобы использовать условия среды, мигрируя через отверстия диаметром всего в несколько микронов, плазмодии могут проникать через крошечные отверстия в разлагающейся древесине. Некоторые плазмодии требуют большего количества влаги, чем другие. Тип афаноплазмодия проводит большую часть своего существования в пределах подложки, в отличие от фанероплазмодия, который лучше приспособлен к жизни на поверхности подложки. Фанероплазмодий имеет хорошо развитую слизистую оболочку, которая служит, чтобы защитить его от потери воды; афаноплазмодий не имеет такой оболочки (Alexopoulos 1960).
2.5.2. Связи с субстратом
Хотя плазмодий может мигрировать далеко от его исходного субстрата, наблюдения, и сбор указывают, что некоторые виды миксомицетов имеет тенденцию закреплятся за определенными типами подложек (Pratt 1934, Gray и Alexopolous 1968, и Stephenson 1988). Некоторые виды почти всегда обитают на коре или древесине, а другие на опаде и разлагающихся растениях. Причины для специфики выбора субстрата неизвестны (Eliasson 1981, Blackwell 1984), однако, они могут, вероятно, быть приписаны физическому и биотическому факторам (Stephenson 1988).
2.5.3. Сезон плодоношения
Сезон образования спорофоров для большинства видов миксомицетов в умеренных областях(регионах) начинается в начале лета и продолжается всю осень. В то время как некоторые виды образуют споры в течение полного периода, другие плодоносят сезонно. На востоке Северной Америке, Lycogala epidendrum может быть найдена с раннего лета до поздней осени.
Ceratiomyxa fruticulosais –плодоносит летом и не встречается осенью. Metatrichia vesparium –изредко встречается и осенью (Stephenson с 1988). В умеренных областях, myxomycetes не развиваются в течение зимних месяцев, хотя некоторые виды могут появляться в течение периодов оттепелей. В тропических районах, myxomycetes остаются активными круглый год, но их плодоношение не особенно обильно кроме некоторых периодов (Lazo 1966).
Главные факторы, воздействующие на сезонное распределение - влажность и температура (Alexopoulos 1968). Некоторые разновидности имеют температурные предпочтение. Blackwell и Gilbertson (1984) сообщили, что культуры влажной камеры Physarum straminipes из Пустыни Sonoran юго-запада Соединенных Штатов плодоносили более быстро при 20o C, чем при 30o C. В течение более раннего полевого изучения, выполненного в той же самой области, P. straminipes никогда не были собраны летом; они появлялись в течение зимних месяцев. Температура, кажется, наиболее важный фактор, основанный на данных, представленных коллекциями, сделанными равномерно в течение годичного периода в двух местах изучения в Бразилии (Maimoni-Rodella и Gottsberger 1980). Они нашли, что даже в присутствии адекватной влажности, myxomycetes были редки, когда температуры были ниже 14o C.
Хотя температура и влажность - важные факторы в определении сезонного распределения для myxomycetes с ограниченными плодоносящими периодами, наше знание этого аспекта экологии все еще не полно.
Как показывают проведенные в некоторых районах Среднего и Северного Урала, из тех видов, что предпочитают образовывать плодовые тела на древесине 65% используют также другие субстраты, и лишь 35% могут быть обнаружены только на древесине, причем 84% всех ксилофильных видов обитают как на древесине хвойных, так и лиственных пород, 11% в большей степени приурочены к хвойным, 5% — к лиственным.[4]
2.5.4. Миксомицеты и насекомые
Myxomycetes обеспечивают пищу, убежище, и место размножения для различных разновидностей насекомых. Самые обычные партнеры myxomycetes в умеренных лесах - жуки. Члены семейства Leiodidae неоднократно собирались с плодоносящими тела и плазмодиями. Жуки играют роль в рассеивании спор. Большинство жуков найдено со зрелыми плодоносящими телами, плазмодии - также важный источник продовольствия (Blackwell 1984, Wheeler 1987). Различные виды летающих насекомых - обычные партнеры myxomycetes (Buxton 1954). Разновидности мух, связанных с myxomycetes, принадлежат семействам Mycetophilidae, Scaridae и Drosophilidae. Муха Epicypta testata, наиболее часто находится в сотрудничестве с такой разновидностью как и Lycogala epidendrum. Sellier и Chassain (1976) сообщили, что взрослая муха иногда откладывает яйца около плазмодия Enteridium lycoperdon и Tubifera ferruginosa. Когда появляются личинки, они кормятся на плазмодии до окукливания, которое происходит одновременно со спорообразованием у миксомицетов. Когда взрослая муха появляется, споры прочно удерживаются на ее теле и разносятся ею.[2]
2.5.5. Миксомицеты и грибы
Плодоносящие тела миксомицетов могут служить как органическая подложка для различных разновидностей грибов. Эти грибы - прежде всего hyphomycetes, хотя некоторые - аскомицеты. Многие из hyphomycetes - фактически вегетативные стадии аскомицетов. (Stephenson и Studlar 1985).
2.5.6.Значение для человека
Так как их существование неизвестно большинству людей, myxomycetes имеет небольшое воздействие на человека. Лишь рассмотренные выше плазмодиофоровые могут вредить хозяйственным культурам. Некоторые миксомицеты употребляются в пищу.
2.6.Географическое Распределение
Слизевики наиболее обычны в экосистемах леса, где имеется разнообразные материала подложки типа коры, пней и опавшего листа. Однако, myxomycetes могут находится, где есть растительные останки. Существуют водные виды миксомицетов. Didymium difforme способен к прохождению полного жизненного цикла под водой. Плазмодии наблюдаются в чашечках водных цветков.
Arcyria cinerea, A. denudata, Certatiomyxa fruticulosa, Comatricha typhoides, Lycogala epidendrum, и Stemonitis fusca - некоторые из наиболее часто встречающихся видов.
2.6.1. Myxomycetes, связанный с определенными Средами обитания
2.6.1.1 Миксомицеты, обитающие на коре деревьев
Миксомицеты одни из постоянных обитателей разлагающейся древесины. Их обилие и видовое разнообразие на этом субстрате довольно высоко. Это может объясняться наличием достаточно разнообразного материала для питания — бактерии, высокомолекулярные продукты разложения древесины, споры и гифы грибов, водоросли и др. С другой стороны разлагающаяся древесина способна достаточно долго удерживать влагу, необходимую для жизнедеятельности трофических стадий — миксамеб и плазмодия, температурные колебания внутри древесины относительно небольшие в течении суток, что также немаловажно. Древесина содержит все биогенные элементы, накопленные в течении жизни деревом, необходимые для нормального существования организма. В процессе разложения древесины грибами эти элементы высвобождаются и становятся доступными для потребления. Все это создает довольно благоприятный микроклимат для жизнедеятельности миксомицетов, особенно проявляющийся, как нам кажется на третьей стадии разложения. Еще одной возможной причиной высокого обилия миксомицетов на древесине в целом может являться отсутствие конкуренции со стороны других организмов. Большое количество видов, которое можно обнаружить на одном субстрате свидетельствует также и о низкой конкуренции среди самих миксомицетов или использовании разными видами разных трофических ресурсов, предлагаемых разлагающейся древесиной.[4]
2.6.1.2. Myxomycetes обитающие на листовой подстилке.
Diderma effusum, D. testaceum, Didymium melanospermum, Lamproderma scintillans, Physarum bivalve и P. cinereum склонны к листовой подстилке. Хотя Arcyria cinerea найден на всех типах листьев, Stemonitis herbatica, кажется, предпочитает листву широколиственных деревьев, в то время как Cribraria microcarpa предпочитает хвойный опад.[2]
2.6.1.3. Экскременты как микросреда обитания
Хотя количество данных ограничено, более чем 80 видов копрофильных myxomycetes были зарегистрированы, с несколькими видами, встречающимися исключительно на экскрементах. Их роль в разложении экскрементов к настоящему времени неизвестна. Из-за высокой влажности, обилия пищи богатства и больших популяций микроорганизмов, экскременты создают подходящую среду обитания для myxomycetes.
Относительно необычные в умеренных лесах, копрофильные миксомицеты, кажется, чаще встречаются в прериях.
2.6.1.4.Почва как Микросреда обитания
Thom и Raper (1930), Warcup (1950) и Indira (1968) изолировали миксомицетов от почвы. Раньше, Feest и Madelin (1985, 1988a, 1988b) демонстрировали метод определения плазмодий-образующих частиц в образцах почвы. Myxomycetes - обычные жители большинства почв. Didymium spp. Кажется, наиболее обильный и широко распространенный слизевик в почвах (Feest и Madelin 1988a).
2.7.Структурные характеристики Myxomycetes
Плазмодий. Отличительным признаком миксогастриевых служит свободноживущий многоядерный плазмодий с амебоидным движением. Отсутствие клеточной стенки у плазмодия создает благоприятные условия для фагоцитоза и пиноцитоза и, следовательно, внутриклеточного пищеварения, т.е. голозойного способа питания (Reyter, Chastellier, 1977; Madelin, 1984). Плазмодий может активно мигрировать и обладает различными положительными и отрицательными таксисами (Konijn, Koevenig, 1971; Hader, Schrecken-bach, 1984). Способ движения аналогичен способу передвижения некоторых саркодовых. Из плазмодия миксомицетов выделен сократительный белок актин, участвующий в организации движения (Hatano et, al. , 1980).[3]
На основании наблюдений за плазмодиями разных видов в условиях культуры было выделено 3 основных типа: протоплазмодий, афаноплазмодий, фанероплазмодий (Alexopoulos, 1960). На ранних этапах развития все 3 типа имеют значительное морфологическое сходство, исчезающее на более поздних стадиях. Протоплазмодий имеет микроскопические размеры и характеризуется отсутствием циркулирующих токов протоплазмы. Этот тип плазмодия характерен для представителей порядков Echinosttlialts и Liceales. Однако если протоплазмодии эхиностелиевых не обладают способностью к слиянию ( Haskins,1978), то протоплазмодий видов лициевых способны сливаться друг с другом (Wollman, Alexopoulos, 1967).
Афаноплазмодий характерен для видов порядков Stemonitalеs. Его отличает отсутствие развитого слизевого чехла, имеющегося у других типов плазмодия, а также наличие особой стадии „коралла" перед началом формирования спорофоров; кроме того, этот тип плазмодия менее устойчив к обезвоживанию, чем остальмые ( Collins, 1979).
Фанероплазнодий наиболее обычен для видов порядка Physarales (Rammeloo, 1976), но он имеется и у некоторых видов порядка Licealee (McManus, l966). Этот тип плазмодия окрашен в различные оттенки красного, желтого, розового, коричневого, черного цветов. В последнее время появились сведения о достаточной стабильности этого признака в условиях культуры ( Coll ins, 1079). Однако в целом таксономическая ценность этого признака низкая.
В отличие от афаноплазмодия фанероплазмодий, так же как и протоплазнодий, может обитать в менее влажной среде. Стадия „коралла" также отсутствует, а при образовании спорофоров фанероплазмодий, как и протоплазмодий эхиностелиевых, может претерпевать плазмотомию. Весьма вероятно, что это сходство может быть связано с филогенетической близостью Echinosteliales и Physarales.
В последнее время стали выделять четвертый тип плазмодия — плазмодий видов порядка Trichiales, занимающий по своей морфологии промежуточное положение между афаноплазмодием и фанероплаэмодием. Он характеризуется наличием токов протоплазмы, но отличается от последнего меньшими размерами и слабой пигментацией (Indira, 1964; Rammeloo, 1976). При неблагоприятных условиях для роста и питания плазмодий превращается в склероций.
У некоторых видов с крупным фанероплазмодием склероций сохраняет его форму, у большинства же видов образуются маленькие склероции (макроцисты), так как плазмодий предварительно дробится. Афаноплазмодий и протоплазмодий не образуют склероция, а только комплекс макроцист, покрытый общим чехлом (Aldrich, Blackwell, 1976). Стенки склероция, так же как и слизевой чехол плазмодия, состоят из фибриллярного матрикса, упакованного более плотно в склероции и менее — в плазмодии. Однако слизевой чехол состоит в основном из галактозы, тогда как стенки склероция — преимущественно из галактозамина (McCormic et al. , 1970).
Для миксогастриевых наряду с наличием многоядерного плазмодия характерен особый тип спороношения, при котором плазмодий превращается в спорофор. Структуры этой стадии обладают наиболее важными таксономическими признаками. К ним относятся гипоталлус или подслоек, который образуется как основание спорофора; перидий, покрывающий споровую массу и имеющий вид либо тонкой, прозрачной или непрозрачной мембраны, либо плотной оболочки, он может быть одно- или многослойным; колонка (колумелла) — стерильная часть спорангия, может быть непосредственным продолжением ножки или дериватом нижней части спорофора; ложная колонка (псевдоколумелла ) — агрегат извести или узелков капиллиция в плоскости спорофора; капиллиций — система нитей, трубочек, пленок, служащая для рассеивания спор. Кроме этих признаков в таксономии миксогастриевых используется еще ряд признаков структур спорофоров, подробный анализ которых будет дан ниже.
Из спор, сформировавшихся в полости спорофора, выходят зооспоры или миксамебы . Эти две формы одной и той жe стадии способны переходить одна в другую путем редукции или образования жгутика. Обычно этот процесс происходит при переходе зоопор из жидкой в более сухую среду и наоборот. Как миксамебы, так и зооспоры способны к амебоидному движению и фагоцитозу. Ползущая зооспора имеет заднюю псевдоподию и периодически образует передний цитостом, в который длинным жгутом „загоняет" бактерии, поступающие затем в пищеварительную вакуоль. Кроме того, зооспора может передвигаться как типичный жгутиконосец. При неблагоприятных условиях зооспоры и миксамебы инцистируются, образуя микроцисты. Их роль примерно та же, что и у конидий некоторых грибов. В условиях лабораторной культуры можно индуцировать переход в микроцисты, переводя их на среду бедную или лишенную аминокислот, некоторых солей, удлинняя или, наоборот, уменьшая световой режим и т.д. (Madelin. 1984). Зооспоры сливаются и образуют зиготу, из которой формируется плазмодий. Известны 2 варианта жизненного цикла: гетероталлический и апомиктический. Наличие гомоталлизма пока с очевидностью не подтверждено ( Collins,1979).
Сведения о строении, биологии отдельных стадий жизненного цикла приводятся в ряде обзоров и монографий (Gray, Alexopoulos, 1968;Collins, 1979;Goodman, 1980; Sauer, 1982; Madelin. 1984).
Таксономия миксомицетов основывается преимущественно на признаках окончательно сформировавшихся спорофоров, так как систематик имеет дело именно с этой стадией как в поле, так и при изучении гербарных образцов.
Тип спорофоров. Спорофоры миксогастриевых отличаются большим разнообразием. При переходе в генеративную фазу плазмодий превращается в спорофор одного из четырех типов: плазмодиокарп, эталий, псевдоэталий, спорангий. Встречаются виды, обладающие спорофорами промежуточного строения, а также спорофорами нескольких типов в одной колонии.
Плазнодиокарп — наиболее простой тип спорофора , когда весь плазмодий покрывается оболочкой и, не меняя формы, целиком преобразуется в спорофор. Вследствие этого плазмодиокарпы всегда сидячие, среди них встречаются ветвящиеся, сетчатые, повторяющие форму плазмодия в период превращения в спорофор. Плазмодиокарпы встречаются у представителей порядков Liceales, Trichiales, Physarales и не найдены среди Stemo-nitales, Echinosteliales.
При образовании спорангия плазмодий обычно распадается на множество частей, каждая из которых развивается в отдельный спорангий характерной формы, цвета и структуры. Этот тип спорофора встречается во всех порядках и семействах класса.
Эталии — подушковидные образования, формирующиеся из ветвящихся плазмодиев, у которых внутренние ответвления образуют спорангии, в то время как протоплазма внешних (наружных) ответвлений разрушается и преобразуется в кортекс, окружающий весь комплекс спорофоров. Остатки боковых стенок спорангиев иногда сохраняются в виде псевдокапилли-ция.
Характерно, что виды, обладающие эталиями, как правило, никогда не образуют других типов спорофоров. Эталии встречаются только у видов порядков Liceales. Physarales, Stemonitales и не встречаются в порядках Trichiales и Echinosteliales.
Псевдоэталии состоят из множества спорангиев, плотно прилегающих друг к другу, но без потери индивидуальности благодаря сохранившимся боковым стенкам, по крайней мере на ранних стадиях развития. Псевдоэталии могут быть, так же как и спорангии, сидячими или на ножках и встречаются в порядках Liceales, Physarales, реже Trichiales, Stemonitales.
Таким образом, если рассматривать класс в целом, то на этом уровне тип спорофора образует непрерывную гамму переходов от одной формы к другой. Наибольшим разнообразием в этом смысле обладает порядок Physarales, где встречаются практически все типы спорофоров. Однако в некоторых случаях этот признак достаточно стабилен на уровне семейств. Так, представители семейства Reticulariaсеае (пор. Liceales) имеют преимущественно эталии, а виды Liceaceae этого же порядка — спорангии и редко плазмодиокарпы.
Форма спорангия — признак, характеризующийся большим разнообразием. Спорангии могут быть сидячими или на ножке, шаровидными, цилиндрическими, подушковидными, яйцевидными, дисковидными, много-лопастными, сжатыми с боков, конусовидными и т.д. В пределах каждой из перечисленных основных форм можно выделить множество отклонений. Например, разветвленные спорангии отличаются различной формой ветвления, сферические спорангии часто бывают несколько вытянуты или приплюснуты, что придает спорангию в целом другой вид. Форма спорангия, несмотря на сильную изменчивость, даже в пределах вида — признак с высоким таксономическим весом. Он имеет ограниченный предел варьирования в рамках одного вида и может быть использован как критерий видового уровня в сочетании с другими признаками.
Окраска спорофора. Значение этого признака в пределах высших таксонов различно. Для примера можно сравнить семейства Triсhiaceae(Trichiales), Stemonitaceae (Ctemonitales), Physaraceae (Physrales). В сем. Trichiaceae преобладает красная, желтая и коричневая окраска. Красный цвет различных оттенков встречается наиболее часто у видов р.Arcyria, у одного вида Тгichiа —— Т. Floriformis и у монотипного рода Metatrichia (M. vesparium). Желтый цвет преобладает в родах Тrichia, Hemitrichia. Для сем. Trichiaceae цвет спорофора имеет только видовое значение, причем в пределах одного вида этот признак варьирует незначительно.
В сем. Stemonitaceae преобладает черный и коричневый цвет спорофора, что может служить как родовым, так и видовым признаком. Например, для представителей р. Lamproderma характерен металлический или радужный оттенок в окраске спорангия, а для видов р. Comatricha и Stemonitis —— коричневый.
В сем. Physaraceae цвет — сильно варьирующий признак даже на уровне вида. Это связано прежде всего с различием в количестве и распределении извести по поверхности перидия, что в свою очередь зависит от условий окружающей среды. Так, например, у широко распространенного Physaraum viride окраска спорангия зависит от качества откладываемых крупинок извести. Окраска может быть оранжево-красной , желтой, зеленоватой или даже серой с металлическим блеском. Эта гамма переходов может быть представлена у спорангиев одной колонии. Таким образом, цвет спорофоров — видовой диагностический признак, который можно использовать только с учетом всех возможных вариаций, количество которых неодинаково в разных таксонах. Из-за сильной изменчивости этот признак можно применять только совместно с другими признаками на уровне вида.
Морфогенез, структура, окраска и форма ножки спорофора. На основании наблюдений за развитием ножки спорофора некоторые авторы разделяют мик-согастриевых на 2 подкласса: Stemonitomycetidae и Myxogastromycetidae (Ross, 1957, 1973). Для первого характерен эпигипоталлический тип развития спорофора, а для второго — субгипоталлический. При эпигипоталлическом развитии ножка — вырост гипоталлуса, а при субгипоталлическом она формируется из тех же участков протоплазмы плазмодия, что и перидий. Если морфогенез ножки действительно протекает двумя различными путями, то это — признак большого таксономического веса, и с его помощью можно решать вопрос о месте в системе даже такого рода с неясным положением, как Diachea. Однако для окончательного решения необходимы дальнейшие исследования особенно на ультраструктурном уровне.
Структура ножки — хороший признак на уровне рода. Например, у видов Comatricha она заполнена фибриллярным материалом, у Macbfideola — полая, а у Stemonitis — полая или заполнена аморфным материалом. В порядке Trichiales этот признак может иметь значение на видовом уровне. Окраска ножки может иметь значение как на уровне рода, так и вида. Форма ножки, а также ее отсутствие или наличие, может служить диагностическим признаком на уровне рода или вида, хотя из-за вариабельности к этому признаку надо относиться с осторожностью. Например, у Trichia varia имеется вся гамма переходов от спорангиев с хорошо выраженной ножкой до сидячих.
Тип растрескивания спорофора при созревании. Наиболее простой способ раскрытия спорофора выражается в неупорядоченном растрескивании перидия. Этот способ часто встречается у видов с плазмодиокарпом и значительно реже у видов со спорангиями. Более специализированный тип растрескивания — разрушение только апикального конца спорофора, при этом растрескивание может осуществляться продольной щелью (Licea biforis), кольцевой щелью (L. parasitica), кольцевой щелью с образованием крышечки (L. kleistobolus). Наконец, у некоторых видов Licea растрескивание происходит вдоль специальных швов на перидии. Роды, у которых способ растрескивания перидия является одним из объединяющих признаков, отличаются гомогенностью своего состава: например, Cribraria, Dictydium. в Cribrariaceae, Dictydiaethalium в Reticulariaceae, Craterium в Physarасеae и т.д. Следовательно, этот признак может быть ключевым на уровне рода, так как он отличается малой изменчивостью и легко наблюдается у гербаризированных образцов. В некоторых случаях, например для видов Licea, он может быть диагностическим на видовом уровне.
Тип капиллиция. Настоящий капиллиций присутствует во всех порядках эндоспоровых миксомицетов. Он образуется как система вакуолей различной формы в протоплазме плазмодия в период превращения последнего в спорофор ( Gray, Alexopouios, 1968; Ellis et al., 1973; Keller et al., 1973; Gaither, 1974). У некоторых родов стемонитовых Comatricha, Stemonitis капиллиций формируется путем образования нитей, отходящих от ножки или колонки (Ross, 1973). Псевдокапиллиций встречается у большинства видов со спорофорани в виде эталиев или псевдоэталиев и, как правило, является остатком перидия боковых стенок редуцированных спорангиев. Так, псев-докапиллиций имеется у видов сем. Reticulariaceae (Liceales) , а также у Fuligo, Мuсilаgо ( Pbysarales). Псевдокапиллиций Reticulariaceae представлен тяжами или перфорированными пластинками (Reticularia), скульптурированныни или гладкими трубочками неправильной формы (Lycogala), щетинками (Тubifеra) или утолщенными частями боковых стенок спорангия , которые свешиваются с краев апикальных концов спорангиев в виде нитей и образуют каркас псевдоэталия (Dictydiaetalium). Сеточки Dictydium и Cribraria, очевидно, гомологичны псевдокапиллицию указанных родов.
Принято считать, что эволюция капиллиция и псевдокапиллиция шла в направлении интенсификации функции рассеивания спор путем усложнения структуры, .т.е. a priori предполагается селективное значение этого признака (Ячевский, 1907; Martin, Alexopoulos, 1969). Однако нам кажется, что этот вопрос может быть решен только после специальных исследований. Пока же следует отметить, что среди Licea, Perichaena, Echinostelium и некоторых других родов имеются виды с частично редуцированным или полностью исчезнувшим капиллицием, причем, несмотря на это, споры прекрасно распространяются после созревания. Изучение структуры капиллиция привело к изменению взглядов на положение некоторых родов в системе. Так, электронно-микроскопические исследования показали, что капиллиций рода Minakatella полый, вследствие чего этот род логично перевести из Dianemaceae в Trichiaceae (Keller et а1., 1973), так же обстоит дело и с родом Ргоtotrichia (Ellis et а1. , 1973). У стемонитовых форма сети капиллиция, особенно его переферической части, может служить ключевым признаком на уровне рода для Comatricha и Stemonitis или видовым для большинства представителей родов Lamproderma и Diachea. Признаки орнаментации капиллиция часто играют важную роль как критерий видового уровня в сем. Trichiaceae, так же как и узелки с известью капиллиция в сем. Physaraceae.
Соли и другие включения в структурах спорофора. Среди миксомицетов широко распространены механизмы выведения кальция в виде солей на стадии перехода плазмодия в спорофор. В частности, углекислый кальций откладывается в ножке спорангия, в капиллиции, в перидии или на нем у видов Physarales. По своей структуре известь может быть либо в форме аморфных гранул ( представители сем. Physaraceae) , либо в виде кристаллов (Didymium) или даже в форме конгломератов кристаллов (Lepidoderma). Из других таксонов кальций найден в спорофорах некоторых видов Trichia contorta, Calomyxa metallica, Metatrichia vesparium —— у последнего в составе органического соединения ( Nelson et al., 1977). Из Других порядков кальций обнаружен в ножке спорангия у Diachea leucоpodia и в перидии основания спорангия Lepidoderma iridescens ( Stemonitales) , а также в диктидиновых гранулах Сribrаriа (Liceales) (Schoknecht, 1975). Однако следует отметить, что у Реrichaena кальций присутствует в форме оксалата, тогда как у представителей Physarales —— в виде углекислого кальция и фосфата. Кроме того, кальций у видов Physarales концентрируется в основном между мембранами перидия, а у Реrichaenа — на его поверхности. Кроме кальция в состав отложений у видов Реrichaena входит кремний.
Таким образом, отсутствие или наличие извести, ее распределение в спорофоре, а также тип известковых отложений позволяют судить о границах между семействами, порядками и даже родами. Крайне необходимы тщательные исследования процессов биоминерализации спорофоров, в частности с помощью метода спектрометрического анализа совместно со сканирующей электронной микроскопией (СЭМ), для установления родственных отношений некоторых родов. Цвет, количество и распределение извести могут сильно варьировать в пределах одного вида, поэтому этот признак на видовом уровне используют в комплексе с другими.
В систематике некоторых групп используются такие признаки, как отложения в виде маслянистого вещества в колонке, стенке спорангия, капиллиция, что характерно для рода Eloеomyxa (Stemonitales). Наличие диктидиновых гранул определяет сем. Cribrariaceae (Liceales).
Перидий. Пленчатый, необызвествленный, прозрачный перидий встречается у видов Licea. и у большинства родов Trichiaceae. а также в некоторых родах Stemonitaceae. Систематическое значение в качестве родовых признаков имеют только отклонения от этого типа, причем наиболее существенны особенности структуры перидия в конце созревания, когда происходит растрескивание спорофора, а также отложение солей или диктидиновых гранул в перидии. Количество слоев также может быть сходным у родов, далеко отстоящих друг от друга. Двуслойный перидий встречается у родов Perchaena (Trichiales) , Physarum и Diderma (Physarales) . В данном случае это может быть результатом параллельной эволюции. Строение поверхности перидия в комплексе с другими данными (наличие извести, тип растрескивания) может играть роль родового признака, поэтому особый интерес могут представлять исследования с применением СЭМ сканирующего электронного микроскопа. Примером такого рода работ могут служить иследования комплекса видов Trichia (Rammeloo, 1974).
Колонка (колумелла), как и ножка, развивается из разных зачатков у стемонитовых и у остальных миксогастриевых. Это обстоятельство необходимо учитывать при анализе признака. Строение и взаимоотношение колонки с системой капиллиция можно использовать как критерий на родовом и видовом уровне, особенно в сем. Stemonitaceae. В пределах Physarales колонка чаще всего встречается у Didermа, Didimium, Lepidoderma и является выростом основания спорангия.
Ложноколонка (псевдоколумелла) — особое образование, встречающееся у некоторых видов Craterium и Physarum, представляет собой массу обызвествленных нитей капиллиция, собранных в сферическую или неправильной формы структуру. Этот признак может сильно варьировать в пределах данного вида и поэтому имеет низкую таксономическую ценность. Однако его следует учитывать в диагнозах видов.
Подслоек (гипоталлус). Эта структура образуется в основании спорофора в виде тонкой пленки или губчатой, роговидной или обызвествленной мембраны. Окраска гипоталлуса, а также его структура в некоторых родах — видовой признак.
Форма, окраска и орнаментация спор. У большинства миксомицетов споры сферические, реже яйцевидные, эллипсоидальные. Споры бывают свободными или объединенными в комплексы, в последнем случае они обычно полиэдрической или неправильной Формы. Форма спор часто нарушается при подготовке постоянных препаратов для световой и электронной микроскопии (так называемый коллапс спор), поэтому желательно проводить обезвоживание спор постепенно и в соответствии с принятыми методиками. Форма и размер спор недоразвившихся спорофоров, а также спорофоров, сформировавшихся в неблагоприятных условиях, также могут сильно отличаться от нормы. Цвет споровой массы и отдельных спор — один из важнейших диагностических признаков. Окраска спор в массе часто не совпадает с цветом отдельных спор в проходящем свете, наблюдаемым на препаратах под микроскопом. Указанные признаки у некоторых видов могут сильно варьировать. У видов р. Echinostelium цвет споровой массы белый, бежевый, розоватый. Под микроскопом споры выглядят практически бесцветными (прозрачными). У некоторых видов, например р. Licea, толщина оболочки неравномерная, из-за чего некоторая часть (обычно примерно треть) выглядит более светлой, чем остальная.
Структура экзоспория у миксомицетов имеет важное значение при разграничении видов. В большинстве монографий и определителей используются данные о строении экзоспория, полученные с применением иммерсии. В последнее время появились работы по изучению тонкой структуры экзоспория методом СЭМ (Schoknecht, Small,1972; Rammeloo, 1974, 1975;1990). Однако, на наш взгляд, в них отсутствует строгое определение элементов структуры экзоспория, что усложняет использование этих признаков для диагностики видов. В этом отношении весьма удобной оказалась классификация типов экзоспория спор головневых грибов (Каратыгин, Азбукина, 1989).Основными элементами структур экзоспория являются бородавочка и шип. Основное различие этих структур в том, что бородавочка — структура с закругленной вершиной, а шип имеет заостренную вершину. Бородавочки могут быть одно- или многовершинными или булавовидными. В последнем случае диаметр вершины больше диаметра основания.
Гладкий тип экэоспория довольно редко встречается у спор миксомицетов, чаще всего этот тип представлен у видов порядков Liceales и Echinosteliales и практически не известен среди Physaralеs.
Бородавчатый и шиповатый тип экзоспория наиболее часто встречается у видов порядков Physarales и Stemonitales. В том случае, если бородавочки или шипы можно различить только при увеличении свыше х900, тип экзоспория можно условно отнести к мелкобородавчатому или мелкошиповатому соответственно. В ряде случаев, если размер бородавок или шипиков явно больше 0.8 мкм, тип спор можно отнести к крупнобородавчатому или крупношиповатому . Как бородавки, так и шипы могут распределяться на поверхности спор относительно равномерно или образовывать скопления .
Сетчатый тип экзоспория широко распространен среди видов порядка Trichiales , Stemonitales , Physarales . Этот тип орнаментации имеет несколько модификаций: сетчато-бородавчатый — с ребрами ячеек, образованными слившимися бородавочками, как например у Comatricha irregularis , Lamproderma atrosporum , сетчато-складчатый — с ребрами уплощенными настолько, что становятся сходными со складками, как например у Diahema depressum или Protophysarum phloiogenum . Следует отметить, что подобная дифференциация сетчатых типов экзоспория возможна только после изучения в СЭМ, в световом микроскопе даже при использовании иммерсионного объектива эти типы экзоспория, как правило, не различимы.
3. Ученые, работавшие в области миксомикологии.
Генрих Антон Бари (1831-1888). Антон Бари признается как основатель современной микологии и первый серьезный миксомиколог. Он рассмотрел Mycetezoa, поскольку он считал их, более близко связанными с protozoans, чем грибы. Его работа Die Mycetezoen была издана в 1859.
Артур Листер (1830-1908)
Джулиельма Листер (1860-1949)
Листеры, отец и дочь, были любителями-натуралистами. Их работа очень повлияла на таксономию миксомицетов. Они написали много публикаций, обращая особое внимание на окружающие условия и их действия на плодоносящую структуру и ее возникновение. «Monograph of the Mycetezoa» была впервые издана в 1894.
Обученая прежде всего дома, Джулиельма Листер была неплохим художником и натуралистом. Еще до появления «женских прав» она была почетным членом Британского Микологического Общества (в 1912 и 1932 президентом).
Томас Хустон Макбрид (1848-1934)
Томас Макбрид заинтересовался миксомицетами через интерес к ботанике. Он был ответственен за увеличение гербария в Государственном университете Штата Айова, включая коллекции myxomycetes со всех континентов. Его первая работа по myxomycetes была монография, «Миксомицеты востока Штата Айова», изданная в 1891. В 1899 была издана книга «Североамериканские слизевики». Незадолго перед его смертью была издана книга «The Myxomycetes», написанная в сотрудничестве с G. W. Мартином. Из известных тогда видов 64 были описаны и названы Макбридом и Мартином.
Джордж Эдвард Масси (1850-1917)
Монография Масси была основана главным образом на коллекциях, размещенных в гербарии Kew. Хотя она все еще используется для номенклатуры и систематики, многие виды, включенной в монографию Масси оказались синонимичны.
Джозеф Томас Ростафинский (1850-1925)
Работа Ростафинского вводила использование микроскопических характеристик для идентификации миксомицетов. До этого, идентификация была ограничена макроскопическими различиями в морфологии. Работы Ростафинскоо, включая «Versuch eines Systems der Mycetezoen» (1873) и «Sluzowce Monographia» (1874, 1875) изданы в Польше.
II. Результаты практических исследований
1. Постановка эксперимента
1.1. Описание образцов.
19 октября в лесопарковой зоне г. Екатеринбурга были собраны образцы коры различных деревьев и пронумерованы:
Образцы
1-10 кора сосны
11 – осина
12, 13 – тополь
14 – ель
15 – ива
16 – груша
17 – черемуха
18 – ясень
19 – рябина
20 - липа.
1.2. Методика эксперимента
19 октября 2000. Были приготовлены влажные камеры (п. 2.4.). В стеклянные чашки Петри была помещена фильтровальная бумага, а поверх бумаги вышеуказанные образцы. Чашки Петри были пронумерованы согласно номерам образцов. В чашки была залита дистиллированная вода, так, что образцы были закрыты водой примерно до середины высоты. Образцы были оставлены при комнатной температуре и рассеянном свете. Через сутки вода была слита. Был проведен первый осмотр при помощи бинокулярной лупы МБС-10 при увеличении Х26 или Х56 по необходимости. Просмотры проводились с 20 октября по 6 декабря включительно раз в 1-2 дня.
2. Результаты
В чашках 5, 9, 12, 13, 18 были обнаружены организмы, относящиеся к миксомицетам.
2.1. Определение видов
Определение было произведено при помощи определителя Новожилова [3].( Рис. См. Приложение II ).
В чашке 5 – Arcyria pomiformis
9 – Paradiacheopsis sp.
12 – Arcyria cinerea
13 - Arcyria cinerea
18 – Physarium decipiens
2.2. Характерные особенности обнаруженных видов
Arcyria pomiformis
Спорофоры – спорангии, рассеянные, редко скученные по 2-3, на ножках, редко сидячие, шаровидные, яйцевидные, желтые разных оттенков, светло-коричневые, грязно-зеленые, до 2 мм выс. , 0,3—0,7 мм диам. Гипоталлус слабо выражен, маленький. Перидий сохраняется в виде чашечки. Чашечка блюдцевидная, складчатая, орнаментирована скученными бородавками и слаборазвитой сеточкой. Ножка цилиндрическая, гладкая, светло- или темно-коричневая, 1/3 – 1/2 от общей высоты спорангия. Капиллиций состоит из нитей, образующих сеточку, прочно связанных с чашечкой. Сеточка капиллиция эластичная, вытягивающаяся как в длину, так и в ширину. Нити капиллиция орнаментированы полукольцами, шипиками, часто имеют расширенные окончания, светло-желтые в проходящем свете, 2—8 мкм диам. Споры в массе охряные, желтые разных оттенков.
Споры 6(7)—8(10) мкм диам., шаровидные, одиночные, бесцветные, бородавчатые или гладкие; бородавки сгруппированы в комплексы.
Плазмодий белый.
На гниющих растительных остатках, на коре живых растений, преимущественно лиственных.
Paradiacheopsis sp.
Спорофоры — спорангии на ножках, шаровидные. Ножка расширяющаяся книзу, заполненная фибриллами, особенно в нижней части. Перидий разрушается после созревания спорангия, иногда сохраняется в виде отдельных пленок. Колонка не достигает середины спороносной части спорангия, ветвится на нити капиллиция. Нити дихотомически ветвятся, никогда не образуют замкнутую сеть.
Arcyria cinerea
Спорофоры — спорангии, скученные или рассеянные, на ножках, полусферические, яйцевидные, обратно яйцевидные, серые, 0.1—0.8 мм в диаметре, 0.3—4.0 мм в высоту. Перидий после созревания спорангия сохраняется в виде чашечки, иногда в виде отдельных фрагментов в апикальной и боковых частях спорангия. Чашечка маленькая, неглубокая, гладкая или орнаментирована очень мелкими бородавками. Ножка тонкая, цилиндрическая, темная или светлая, 0.2—2 мм выс. , иногда общая для нескольких спорангиев, заполнена спороподобными клетками 15—21 мкм диам. Капиллиций состоит из нитей, формирующих сеточку. Нити капиллиция крепятся к внутренней поверхности чашечки, бесцветные или светло-желтые в проходящем свете; во внутренней части сети нити гладкие и толстые, в средней части тонкие и с шипиками, в наружной — с более длинными шипиками, бородавками и полукольцами. Споры в массе серые или желто-серые.
Споры 6—7 мкм диам., шаровидные, одиночные, бесцветные, бородавчатые.
Плазмодий белый, серый, серо-желтый.
На гнилых растительных остатках, коре живых деревьев, преимущественно хвойных, на помете растительноядных животных.
Physarium decipiens
Спорофоры — спорангии или маленькие плазмодиокарпы (в моем случае серебристый плазмодиокарп), скученные, сидячие, полусферические, подушковидные. Желтые или оранжевые, 0.3—0.7 мм диам. , до 2 мм дл. Гипоталлус пленчатый, коричневый. Перидий однослойный, пленчатый, равномерно инкрустирован известью в виде бугорков и отдельных чешуек. Капиллиций состоит из маленьких, удлиненных, звездчатых, желтых или красных узелков с известью, соединенных прозрачными трубочками. Споры в массе черные,
Споры 9—14 мкм диам., темно-коричневые, мелкошиповатые.
Плазмодий желтый.
На гнилых растительных остатках, на коре живых деревьев.
3. Выводы
Полученные результаты согласуются с данными [2], изложенными в п.2.4.1.
Миксомицеты обнаружены на 5 из 20 взятых образцов коры. Причем из 11 образцов коры хвойных деревьев (10 образцов коры сосны и один образец коры ели ) миксомицеты проросли на двух.
Из 9 образцов лиственных деревьев миксомицеты обнаружены на трех. Причем на образцах с гладкой корой ( осина, ива, груша, черемуха, рябина, липа ) миксомицеты вообще не обнаружены, а на всех образцах с грубой корой ( тополь-2 образца, ясень ) миксомицеты проросли.
Исходя из результатов эксперимента также можно сделать вывод, что наиболее распространены в лесопарковой зоне г.Екатеринбурга миксомицеты рода Arcyria.
СПИСОК ЛИТЕРАТУРЫ
Курс низших растений. Под общей редакцией М.В.Горленко, М., Высшая школа, 1981
Myxomycetes: A Handbook of Slime Molds. Steven L. Stephenson. Published by Timber Press.
Определитель грибов России. Отдел myxomycota, вып. 1. Класс Myxomycetes. Ю.К. Новожилов, Санкт-Петербург, Наука, 1993.
Экология процессов биологического разложения древесины. Российская академия наук Уральское отделение Институт экологии растений и животных.изд. «Екатеринбург» 2000.
State indicators. State and change of Biodiversity. Russian A c ademy of Sciences. Institute for the history of science and technology.
Протозоология. К. Хаусман. М. «Мир» 1988
«Мир растений» под общ. ред. чл.-корр. АН СССР М.В. Горленко
| Видовой состав трутовиковых грибов окрестностей г. Чернигова | |
|
Содержание Вступление Раздел 1. Общие сведения о трутовиковых грибах 1.1 Классификация трутовиков 1.2 Морфология трутовиковых грибов Раздел 2 ... Проникновению спор гриба внутрь древесины препятствует возникновение раневого ядра. Древесные породы заражаются мицелием, развивающимся из попадающих на поверхность коры спор грибов, и проникающим внутрь. |
Раздел: Рефераты по биологии Тип: курсовая работа |
| Биология | |
|
ПРОГРАММА вступительного экзамена по общей биологии для направления "Биология" (магистерская программа "Биология") 1.БИОЛОГИЧЕСКОЕ МНОГООБРАЗИЕ 1.1 ... ... и биологии в связи с паразитизмом, жизненные циклы, основные представители (дизентерийная амеба, лямблия, трипаносомы, лейшмании, кокцидии, токсоплазма, малярийный плазмодий) СЛИЗЕВИКИ / МИКСОМИЦЕТЫ. Строение эукариотной клетки грибов. |
Раздел: Рефераты по биологии Тип: учебное пособие |
| Шпоры по ботанике. Грибы | |
|
28 Общая характеристика грибов . Отличительные признаки. Эволюция мицелия. Это слоевищные безпластидные организмы, имеющие признаки растений и ... Зооспора(с одним жгутиком) -> зарожение кл. хозяина-> проростание тротопласта паразита в клетке хозяина ( видна зооспора пораз. на поверхности клетки хозяина. -> Увеличение таллома ... Вегетативное тело слизевиков - плазмодием это многоядерная цитоплазматическая масса, не дифференцированная на клетки, лишенная какой-либо постоянной формы , микроскопически мал ... |
Раздел: Рефераты по ботанике и сельскому хозяйству Тип: реферат |
| Билеты по биологии для 10-11 классов | |
|
Билеты по биологии БИЛЕТ №1 ВОПРОС 1. Уровни организации живой материи Молекулярный. Любая живая система, как бы сложно она ни была организована ... Спорообразование встречается у простейших (малярийный плазмодий), грибов, водорослей и лишайников. Водоросли снабжают гриб сахарами и получают от гриба минеральные соли, которые тот извлекает из древесины, породы" почвы и др. |
Раздел: Биология и химия Тип: шпаргалка |
| Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних ... | |
|
Реферат На тему: Риниофиты. Бесполое размножение. Спорангии и спорогенез у древних высших растений Риниофиты Риниофиты (Rhyniophyta), псилофиты ... В периплаэмодиальном тапетуме стенки его клеток, иногда уже на ранних стадиях развития спорангия, расплываются к их протопласты сливаются вместе в одну сплошную массу, имеющую ... Эволюция последнего, кроме того, выразилась в создании длинной и тонкой ножки, выработке кольца и в общем уменьшении количества производимых спорангием спор. |
Раздел: Рефераты по биологии Тип: реферат |