Доклад: Проблема происхождения хордовых
В последние годы китайские палеонтологи находят ископаемые организмы поразительной сохранности - даже с отпечатками структур, образованных мягкими тканями. Обнаружены эти остатки в центральной части провинции Юньнань (Южный Китай) в нижнекембрийских глинистых сланцах свиты Маотяньшань, возраст которых около 530 млн лет. Организм, чьи остатки были найдены в 1995 г. в Шэньяне, получил название по имени провинции - юннанозоон (Yunnanozoon) (Chen J.-Y. et al. // Nature. 1995. V.377. P.720-722). Одни специалисты рассматривали его как древнейшего представителя бесчерепных хордовых (Acrania), другие относили эту форму к полухордовым (Hemichordata). А в 1999 г. в Хайкоу, вблизи Куньмина, было раскопано целое кладбище древнейших животных - остатки 305 экземпляров, но другого хордового, названного хайкоуэллой (Haikouella). Благодаря прекрасной сохранности палеонтологам удалось с большой полнотой восстановить ее строение (Chen J.-Y. et al. // Nature. 1999. V.402. P.518-521).
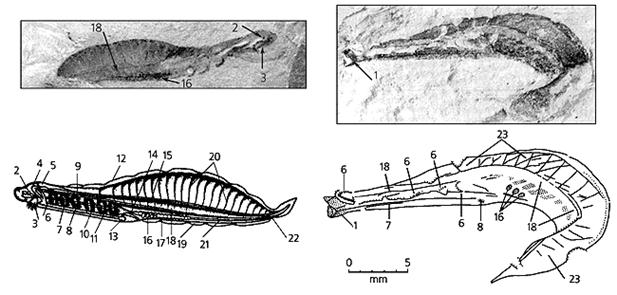
Фоссилизированные остатки (верхний ряд) и реконструкции хайкоуэллы (слева) и юннанозоона. Это самые древние хордовые из когда-либо и где-либо найденных. 1 - передний конец тела, 2 - глаз, 3 - ротовая полость, 4 -передний жаберный сосуд, 5 - мозг, 6 - жаберные щели, 7 - эндостиль, 8 - зубовидные структуры, 9 - спинная аорта, 10 - глотка, 11 - брюшная аорта, 12 - спинной плавник, 13 - сердце, 14 - пищевод, 15 - нервный тяж, 16 - половые железы, 17 - желудок, 18 - хорда, 19 - брюшной плавник, 20 - миосепты, 21 - кишка, 22 - анальное отверстие, 23 - миомеры.
Тело хайкоуэллы было ланцетовидным, длиной 25-40 мм, отчетливо выделялись голова, туловище и хвост. Брюшная часть туловища утолщена, спинная сжата с боков. Имелись плавники: невысокий спинной и небольшие брюшной и хвостовой. По бокам головы расположена пара округлых структур, которые китайские палеонтологи интерпретировали как глаза. На брюшной стороне головы находилось ротовое отверстие, окруженное небольшими щупальцами и ведущее в объемистую глотку, боковые стенки которой пронизывались жаберными щелями и укреплялись шестью парами жаберных дуг. В глотке имелись конические зубовидные выросты, а по ее дну тянулся желобок - эндостиль, - признак, характерный для всех хордовых. Глотка переходила в узкий пищевод, соединявшийся с извитой средней кишкой, за ней следовала прямая задняя, которая заканчивалась анальным отверстием на брюшной стороне тела, у основания хвоста.
Над кишечником хайкоуэллы лежала хорда (осевой скелет), в туловище утолщенная и смещенная в нижнюю часть тела. Над ней располагался продольный нервный тяж, который в голове образовывал мозг из трех отделов. Под глоткой и над ней также находились тяжи, но китайские палеонтологи интерпретировали их как главные кровеносные сосуды - брюшную и спинную аорты. Их связывали жаберные артерии, а сама брюшная аорта начиналась от похожего на пузырь вздутия, которое могло быть сердцем.
У хайкоуэллы спинную часть туловища над хордой занимали метамерные мышцы, сегменты которых (миомеры) разделялись S-образными перегородками - септами. По бокам средней кишки находились половые железы: четыре пары у хайкоуэллы и 13 пар у юннанозоона.
По всем важнейшим диагностическим признакам - общему плану строения тела, наличию хорды, центральной нервной системы в виде продольной трубки, глотки с жаберными щелями - хайкоуэлла и юннанозоон относятся к хордовым животным. До находок в Китае столь древние представители хордовых не были известны, и проблему их происхождения многие поколения ученых пытались решить умозрительно. Интересно, что строение хайкоуэллы и юннанозоона оказалось очень близким к типу организации гипотетического общего предка хордовых, который был обоснован в 1925 г. А.Н.Северцовым и назван им примитивным бесчерепным (Acrania primitiva). Хайкоуэлла отличается от него такими признаками более высокой организации, как обособленная голова, в которой отсутствуют мышечные сегменты-миомеры, головной мозг, парные глаза, сердце. Все это сближает хайкоуэллу с предками высших хордовых - позвоночных.
Полученные китайскими палеонтологами данные соответствуют
гипотезе о происхождении хордовых от древних “олигомерных червей”, близких по
уровню организации к современным кишечнодышащим полухордовым (Enteropneusta).
Вероятно, на ранних этапах эволюции хордовых у их предков развились новые
приспособления (хорда, сегментация туловищных мышц, более совершенные органы
чувств и центральная нервная система), которые обеспечили животным переход от
придонной жизни и червеобразного способа передвижения к активному плаванию в
толще воды. Древнейшие хордовые сохраняли способ питания “олигомерных червей” -
фильтрацию взвешенных в воде микроорганизмов и частиц ила с помощью глоточного
фильтрационного аппарата. Возможно, уже тогда хордовые разделились на три
группы, образовавшие в ходе дальнейшей эволюции три известные ныне подтипа:
оболочников, бесчерепных (головохордовых) и позвоночных (черепных). От
прикрепленных к морскому дну предков произошли оболочники, группа роющих
морских хордовых дала начало бесчерепным, а те, что продолжали совершенствовать
приспособления к активному плаванию, оказались предками позвоночных. Видимо,
хордовые возникли в самом начале кембрийского периода - во время “кембрийской
взрывной эволюции”, когда в палеонтологической летописи появились представители
почти всех известных типов организмов, а может, и еще раньше, в поздневендское
время.
Литература
Статья Н.Н. Иорданского, доктор биологических наук Институт проблем экологии и эволюции им.А.Н.Северцова РАН, Москва