Курсовая работа: Определение активности ферментов
Российский химико-технологический Университет
им. Д.И.Менделеева
Кафедра промышленной биотехнологии
МЕТОДИЧЕСКАЯ РАБОТА
ПО ОПРЕДЕЛЕНИЮ АКТИВНОСТИ ФЕРМЕНТОВ
Выполнила: студентка
группы Э-35
Тимошкина Е.А.
Преподаватель:
Мартимьянова Н.А.
Москва,
1996 г
СОДЕРЖАНИЕ
Свойства ферментов как биологических катализаторов. 1
Получение ферментных препаратов. 4
Общие методы определения активности ферментов. 6
Методы определения активности протеолитических ферментов 11
Литература 19
Свойства ферментов как биологических катализаторов.
Фермент - от лат. fermentum - закваска; знзим - от греч. эн - внутри, зиме - закваска.
Ферменты, или энзимы, - это катализаторы белковой природы, образующиеся и функционирующие во всех живых организмах. Происхождение терминов связано с тем, что первоначально ферментативные процессы были открыты и изучены в бродильном производстве. В каждой клетке имеются сотни различных ферментов. С их помощью осуществляются многие химические реакции, которые могут с большой скоростью идти при температурах, подходящих для данного организма, т.е. в пределах от 5 до 400 С. Чтобы эти реакции с той же скоростью протекали вне организма, потребовались бы высокие температуры и резкие изменения некоторых других условий. Для клетки это означало бы гибель, так как вся работа клетки строится таким образом, чтобы избежать любых сколько-нибудь заметных изменений в нормальных условиях ее существования. Следовательно, ферменты можно определить как биологические катализаторы, т.е. как вещества, ускоряющие реакции. Они абсолютно необходимы, потому что без них реакции в клетках протекали бы слишком медленно и не могли поддерживать жизнь. Совокупность биохимических реакций, катализируемых ферментами, составляет сущность обмена веществ, являющегося отличительной чертой всех живых организмов. Через ферментативный аппарат, регуляцию его активности происходит и регуляция скорости метаболических реакций, их направленности.
Являясь катализаторами, ферменты имеют ряд общих с небиологическими катализаторами свойств:
1. Ферменты не входят в состав конечных продуктов реакции и выходят из нее, как правило, в первоначальном виде, т.е. они не расходуются в процессе катализа (в настоящее время доказано, что некоторые ферменты в конце химической реакции подвергаются модификации и даже распаду, а не освобождаются в неизменном виде, как постулировал Л.Михаэлис).
2. Ферменты не могут возбудить те реакции, протекание которых противоречит законам термодинамики, они ускоряют только те реакции, которые могут протекать и без них.
3. Ферменты не смещают положения равновесия, а лишь ускоряют его достижение.
Специфические свойства:
1. Конечно же, по своему химическому строению все ферменты являются белками.
2. Эффективность ферментов намного выше, чем небиологических катализаторов (скорость протекания реакции при участии фермента выше на несколько порядков).
3. Ферменты обладают узкой специфичностью, избирательностью действия на субстраты, т.е. на вещества, превращение которых они катализируют. Высокая специфичность ферментов обусловлена конформационной и электростатической комплементарностью между молекулами субстрата и фермента и уникальной структурой активного центра фермента, обеспечивающими “узнавание”, высокое сродство и избирательность протекания одной какой-либо реакции из тысячи других химических реакций, осуществляющихся одновременно в живых клетках.
В зависимости от механизма действия различают ферменты с относительной (или групповой) специфичностью и абсолютной специфичностью. Так, для действия некоторых гидролитических ферментов наибольшее значение имеет тип химической связи в молекуле субстрата. Например, пепсин расщепляет белки животного и растительного происхождения, хотя они могут существенно отличаться друг от друга как по химическому строению и аминокислотному составу, так и по физико-химическим свойствам. Однако пепсин не расщепляет углеводы или жиры. Объясняется это тем, что местом действия пепсина является пептидная -СО-NH- связь. Для действия липазы, катализирующей гидролиз жиров на глицерин и жирные кислоты, таким местом является сложноэфирная связь. Аналогичной относительной специфичностью обладают также некоторые внутриклеточные ферменты, например гексокиназа, катализирующая в присутствии АТФ фосфорилирование почти всех гексоз, хотя одновременно в клетках имеются специфические для каждой гексозы ферменты, выполняющие такое же фосфорилирование.
Абсолютной специфичностью действия называют способность фермента катализировать превращение только единственного субстрата. Любые модификации в структуре субстрата делают его недоступными для действия фермента.
Стереохимическая специфичность ферментов обусловлена существованием оптически изомерных L- и D-форм или геометрических (цис- и транс- ) изомеров химических веществ. “Так, известны оксидазы L- и D-аминокислот, хотя в природных белках обнаружены только L-аминокислоты. Каждый из видов оксидаз действует только на свой специфический стереоизомер.
+ЅО2
L-аминокислота ![]() кетокислота
+ NH + H2O
кетокислота
+ NH + H2O
оксидаза L-аминокислот
+ЅО2
D-аминокислота ![]() кетокислота
+ NH + H2O
кетокислота
+ NH + H2O
оксидаза D-аминокислот
Наглядным примером стереохимической специфичности является бактериальная аспартатдекарбоксилаза, катализирующая отщепление СО2 только от L-аспаргиновой кислоты с превращение ее в L-аланин” [1].
4. Регулируемость ферментов как биокатализаторов. “Через регуляцию ферментативного аппарата осуществляется скоординированность всех метаболических процессов во времени и пространстве, направленное на воспроизведение живой метерии, поддержание постоянства внутриклеточной среды, на приспособление к меняющимся внешним условиям” [2].
5. Термолабильность ферментов. Скорость химических реакций зависит от температуры, поэтому катализируемые ферментами реакции также чувствительны к изменениям температуры. Однако вследствие белковой природы фермента тепловая денатурация при повышении температуры будет снижать эффективную концентрацию фермента с соответствующим снижением скорости реакции. Таким образом, термолабильность, или чувствительность к повышению температуры является одним из характерных свойств ферментов, резко отличающих их от неорганических катализаторов. При 1000С почти все ферменты утрачивают свою активность (исключение составляют, очевидно, только один фермент мышечной ткани - миокиназа, которая выдерживает нагревание до 1000С). При низких температурах (00С и ниже) ферменты, как правило, не разрушаются, хотя активность их падает почти до нуля. Во всех случаях имеет значение время воздействия соответствующей температуры. В настоящее время для пепсина, трипсина и ряда других ферментов доказано существование прямой зависимости между скоростью инактивации фермента и степенью денатурации белка. На термолабильность ферментов определенное влияние оказывают концентрация субстрата, рН среды и другие факторы.
6. Зависимость активности ферментов от рН среды. Ферменты обычно наиболее активны в пределах узкой зоны концентрации водородных ионов, соответствующей для животных тканей в основном выработанным в процессе эволюции физиологическим значением рН среды 6.0 - 8.0. рН-оптимум действия ферментов лежит в пределах физиологических значений. Исключение составляет пепсин, рН-оптимум которого равен 2.0. Объясняется это тем, что пепсин входит в состав желудочного сока, содержащего свободную соляную кислоту, которая создает оптимальную кислую среду для действия этого фермента. С другой стороны, рН-оптимум аргиназы лежит в сильно щелочной зоне (около 10.0); такой среды нет в клетках печени, следовательно, in vivo аргиназа функционирует, по-видимому, не в своей оптимальной зоне рН среды. Влияние изменений рН среды на молекулы фермента заключается в воздействии на состояние и степень ионизации кислотных и основных групп (СООН-группы дикарбоновых аминокислот, SH-группы цистеина, имидазольного азота гистидина и др.). При разных значениях рН среды активный центр может находиться в частично ионизированной или в неионизированной форме, что сказывается на третичной структуре белка и соответственно формировании активного фермент-субстратного комплекса. Кроме того, имеет значение и состояние ионизации субстратов и кофакторов.
Получение ферментных препаратов.
Для получения ферментных препаратов используют как микроскопические грибы, так и бактерии и дрожжи. Иногда получение технического ферментного препарата кончается проведением процесса ферментации, например, в спиртовой промышленности для осахаривания крахмала используют жидкую культуру Aspergillus niger. Впоследствии ее добавляют в жидком виде в количестве 10-12% к осахареваемому затору. Однако активность ферментов в культуральной жидкости быстро снижается. Поэтому широко практикуют получение сухих технических ферментных препаратов.
Технические препараты ферментов. Комплексный амилолитический ферментный препарат получают путем выращивания плесневых грибов на твердой питательной среде с последующей сушкой и измельчением полученной массы. Более активный препарат фермента получают путем экстракции такого “грибного солода” с последующим выпариванием и сушкой. Еще более активные ферментные препараты можно выделить из культуральной жидкости путем осаждения амилазы ацетоном и дальнейшим высушиванием коагулятом при температуре 27-270С. Для осаждения фермента часто используют и сульфат аммония. Предварительно культуральную жидкость выпаривают при температуре 400 С до 40%-ного содержания сухих веществ. Коагулят сушат вместе с наполнителем.
Комплекс ферментов протеолитического и амилолитического действия получают при помощи культуры Bacillus subtilis. Это аэробные, грамположительные, подвижные палочки. Для этих бактерий характерен очень богатый комплекс гидролитических ферментов. В качестве источников питания они могут использовать белки, углеводы, спирты, органические кислоты. Bacillus subtilis культивируют как методом поверхностного культивирования на отрубях, так и в жидких средах особого состава по методу глубинного культивирования.
Целлюлолитические ферментные препараты. Производство целлюлаз основывается на использовании культуры гриба Trichoderma viride. Существующие в настоящее время способы получения целлюлаз в глубинной культуре предполагает выращивание микроорганизмов-продуцентов целлюлаз на питательной среде, содержащей в качестве источников углерода, как правило, очищенную целлюлозу, или же содержащие ее природные субстраты. Но получение целлюлазы с использованием в качестве основного компонента среды природной целлюлозы (например, древесные опилки) сопряжено с рядом технологических трудностей. Более рационально использование питательной среды, содержащей растворимый “индуктор”. Такой питательной средой может быть молочная сыворотка, основным компонентом которой является лактоза (предварительно от молочной сыворотки отделяют белок). В качестве продуцента может быть использован гриб Trichoderma lignorum, позволяющий получить весь комплекс целлюлолитических ферментов, необходимый для расщепления природных целлюлозусодержащих субстратов.
Общие методы определения активности ферментов.
Прежде чем преступить к выделению фермента, необходимо избрать и тщательно отработать метод определения активности, под контролем которого производится выбор наиболее эффективных приемов очистки ферментов, а затем и выполнение последовательных стадий его препаративного получения. Активность фермента меняется при различных условиях реакции и зависит от температуры, рН среды, от концентраций субстратов и кофакторов. Учитывая это, при определении активности фермента на разных стадиях очистки необходимо строго соблюдать одни и те же условия. Желательно не ограничиваться определением активности по одному какому-либо методу. Количество субстрата, превращаемого в условиях теста по определению активности фермента, должно быть пропорционально количеству последнего и времени инкубирования. Если же нет такой пропорциональности, то активность рассчитывают по предварительно построенному калибровочному графику, отражающему зависимость скорости реакции от количества единиц фермента. Когда ход реакции нелинеен во времени, следует определять начальную скорость реакции (по тангенсу угла наклона касательной к начальному отрезку кривой превращения). Для этого легче всего применять такие методы изменения активности, которые позволяют непрерывно следить за ходом превращения во времени: спектрофотометрические методы, потенциометрические, полярографические и т.п. Для измерения скорости ферментативной реакции необходимо выбрать буфер, который не тормозит исследуемую активность и обеспечивает поддержание рН раствора, близкой к оптимальной для данного фермента. Реакцию проводят при температуре, лежащей в большинстве случаев в пределах 25-400С. При исследовании ферментов, требующих присутствия кофакторов (ионов металлов, коферментов и др.), концентрация которых может снижаться по мере очистки фермента, в реакционную смесь следует добавлять недостающие кофакторы, например соли металлов, АТФ, НАДФ и т.п. Также для определения активности ферментов вводят стабилизаторы в состав опытных смесей. Во многих случаях добавление желатина, альбумина и других добавок предотвращает денатурацию ферментного белка.
Спектрофотометрические методы. Спектрофотометрические методы основаны на поглощении света в определенных участках спектра многими соединениями, являющимися активными группами ферментов, субстратами или продуктами реакции. Положение максимума поглощения при определенной длине волны определяется наличием в исследуемом материале определенных групп - аналитических форм. Для измерения спектров используют специальные приборы - спектрофотометры, фотометрические абсорбциометры и др. Этот метод отличается высокой чувствительностью, быстротой определения, малым расходованием фермента и реактивов и позволяет следить за течением реакции во времени. Для этого реакционную смесь помещают в кювету, вставленную в термостатируемый кюветодержатель. Через малый промежуток времени после добавления фермента (или субстрата) и быстрого перемешивания измеряют поглощение при длине волны, характерной для используемого субстрата или конечного продукта, образующегося в данной реакции. С помощью спектрофотометрического метода можно измерять непосредственно концентрацию некоторых ферментов (после достаточной очистки) по величине характерных максимумов поглощения прочно связанных коферментов (простетических групп). Необходимо иметь произвольно выбранную единицу фермента, с помощью которой можно было бы количественно выразить чистоту и активность различных фракций. В большинстве случаев выбор единицы зависит от избираемого метода определения. В случае спектрофотометрического метода такой единицей может служить количество фермента, которое вызывает определенное изменение в оптической плотности за определенное время при данных условиях опыта; если определяется продукт реакции, то единицей будет количество фермента, которое вызывает образование определенного количества вещества в минуту, и т.д. Тогда удельную активность фермента, которая является мерилом чистоты ферментного препарата, выражают числом единиц в 1 мг вещества (белка).
Для целей определения ферментов могут быть использованы не только измерение поглощения света, но также измерения флюоресценции - спектрофлюорометрические методы. Такое определение активности фермента в ряде случаев по чувствительности превосходит спектрофотометрические методы на целый порядок величины. Некоторые коферменты и субстраты обладают сильной флюоресценцией. НАД и НАДФ в восстановленном состоянии имеют сильную флюоресценцию и не флюоресцируют в окисленном состоянии. Поэтому спектрофлюорометрию используют для изучения кинетики и механизма действия никотинамидных и флавиновых ферментов.
Колориметрические (фотометрические) методы. В основе этих методов лежит измерение при помощи визуального или фотоэлектрического колориметра окрашенного продукта, образующегося при взаимодействии субстрата или продукта действия фермента со специфическими реактивами, которые обычно добавляются в фиксированную опытную пробу, взятую после остановки ферментативной реакции. Эти методы весьма разнообразны. Разработаны количественные методы, основанные на биуретовой реакции, при которой состав белка, очевидно не должен сказываться на результатах определения, т.к. эта реакция на пептидные связи, а не на боковые группы в белке. Метод Горнелла и соавторов, основанный на измерении полосы поглощения в области 550-650 нм, используется для определения значительных количеств (1-10 мг) белка в пробе. Предлагаются биуретовые микрометоды, основанные на поглощении ультрафиолетовых лучей медно-белковыми комплексами: они позволяют определять 0.1 - 2.0 мг белка в пробе. Число небелковых веществ, которые могут влиять на определения с помощью биуретовой реакции, невелико, но следует вносить поправки на те вещества, которые присутствуют в высоким концентрациях (соли аммония, сахароза). Наиболее ценными являются те фотометрические методы, которые позволяют следить во времени за ходом ферментативной реакции без ее прекращения по изменению окраски хромогенного субстрата или добавленного индикатора. Метод Фолина и Чиокальто, предложенный для определения количества белка, основан на хромогенной природе некоторых боковых групп аминокислот, а также на присутствии в белках пептидных связей. Этот метод обладает высокой чувствительностью (на пробу достаточно 0.01 - 0.1 мг белка), но многие небелковые вещества мешают определению.
В настоящее время для определения количества белка широко пользуются измерениями интенсивности поглощения света при 280 нм, которое обусловлено присутствием в белке ароматических аминокислот. Количество этих аминокислот в разных белках различно и ,следовательно, интенсивность неодинакова. В кювете толщиной 1 см у раствора, содержащего 1 мг “усредненного” белка в 1 мл, оптическая плотность равна 1 при длине волны 280 нм. Нуклеиновые кислоты поглощают при 280 нм, но можно сделать поправку на их присутствие, проводя измерения и при 260 нм и при 280 нм. Очень важна быстрота измерения активности ферментов. То же относится и к измерению количества сухого остатка или количества белка. Тем самым предпочитают быстрый метод определения белка путем измерения величины поглощения при 280 нм. Выигранное время важнее, за счет того, что удельное поглощение выделяемого белка иногда значительно отличается от средней величины поглощения смеси белков, присутствовавших в исходном материале.
Манометрические методы. Эти методы используются при определении активности фермента в тех случаях, когда в исследуемых реакциях один из компонентов находится в газообразном состоянии. К таким реакциям относится главным образом те, которые связаны с процессами окисления и декарбоксилирования, сопровождающимися поглощением или выделением кислорода и углекислоты, а также реакции, в которых выделение или связывание газа происходит в результате взаимодействия продуктов ферментативного превращения с добавленным в систему реактивом. Наблюдение за ходом реакции во времени проводится в специальных приборах - манометрических аппаратах Варбурга.
Другие методы. Сюда относится обширный ряд методов, включающих поляриметрию, вискозиметрию, потенцио- и кондуктометрические измерения и т.п. Также определение активности можно выполнять, используя методы хроматографии и электрофореза на бумаге. Эти методы высокочувствительны и специфичны, что делает их во многих случаях незаменимыми; они позволяют значительно сократить расход фермента на измерение активности, но не всегда применимы ввиду продолжительности разделения веществ в процессе хроматографии (и электрофореза).
Специальные методы определения активности пепсина и папаина. Пепсин и папаин относятся к одной из наиболее многочисленных групп ферментов - протеолитические ферменты (протеазы). Они принадлежат к классу гидролаз, к подклассу пептидгидролаз и катализируют расщепление белков и полипептидов в соответствии с уравнением :
R1CONHR2 + H2O R1COOH + H2NR2 ,
R1 и R2 - остатки аминокислот, ди- или полипептиды.
Т.е. протеазы катализируют расщепление пептидной связи CONH .
Теперь немного подробнее о пептидгидролазах.
Подкласс пептидгидролаз делится на следующие группы:
1. аминопептидазы (аминоацилпептидгидролазы) катализируют гидролиз полипептидов по месту пептидной связи, находящейся рядом со свободной аминной группой.
действие
R1 фермента R2
| |
H2N C C NH C C
║ ║
2. карбоксипептидазы (пептидиламинокислотные гидролазы) катализируют расщепление полипептидов по месту пептидной связи, находящейся рядом со свободной гидроксильной группой.
R
|
CONHCH
|
действие COOH
фермента
3. дипептидазы (дипептидгидролазы) катализируют гидролитическое расщепление на свободные аминокислоты.
NH2CH2CONHCH2COOH + H2O 2CH2NH2COOH
глицил-глицин гликолол
Дипептидазы расщепляют только такие пептидные связи, по соседству с которыми находятся одновременно свободные карбоксильная и аминная группы. Большое влияние на действие дипептидаз оказывает наличие -водородных атомов, находящихся у атомов углерода, связанных со свободной карбоксильной и аминной группами. При замене этих атомов другими группировками активность ферментов снижается или совсем исчезает.
4. протеиназы - (пептидилгидролазы) гидролизуют непосредственно белок. При этом образуются полипептиды и свободные аминокислоты. К этой подгруппе протеиназ принадлежит пепсин, трипсин, химотрипсин, папаин и др. В процессе действия ферментов на субстраты решающую роль играет структура молекул субстрата (соответствующая часть молекулы субстрата), в которой происходит разрыв. Характер распада белковой молекулы на пептиды и аминокислоты зависит от природы субстрата и внешних условий (рН, температуры и др.). В качестве белковых субстратов используют желатин, гемоглобин, казеин, сывороточный альбумин.
Методы определения активности протеолитических ферментов
Модификационный метод. Модификационный метод с применением субстрата казеина основан на определении скорости ферментативной реакции гидролиза субстрата под действием исследуемых протеолитических ферментов, содержащихся в материале, взятом на анализ. Скорость реакции определяют по количеству образовавшихся аминокислот - тирозина (a-амино-b-оксифенилпропионовая кислота) и триптофана (a-амино-b-индолилпропионовая кислота), которые устанавливают колориметрической реакцией с реактивом Фолина. Этим методом определяют указанные аминокислоты как в свободном, так и в связанном состоянии.
По количеству тирозина и триптофана, содержащемуся в гидролизате, определяют количество превращенного белка в процессе ферментативной реакции (из расчета содержания в белке 6% тирозина и 1.8% триптофана). В основу расчета активности протеолитических ферментов положена зависимость степени гидролиза белка от числа единиц активности фермента, введенных в ферментативную реакцию. На основе этой зависимости составляется график.
Поскольку в данном методе количество аминокислот, характеризующих количество превращаемого белка, определяется по интенсивности окраски реакционных сред - оптической плотности после реакции с реактивом, то в графике для простоты расчета вместо количества превращенного белка ставится пропорциональная ему оптическая плотность раствора. При составлении графика по оси абсцисс откладывают число единиц фермента, введенное в ферментативную реакцию, а по оси ординат - оптическую плотность, полученную после колориметрической реакции с реактивом Фолина.
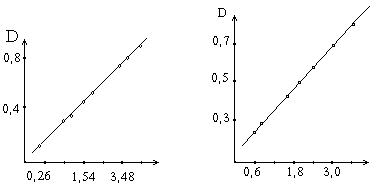
Содержание белка,мг Единицы активности
Зависимость оптической плотности от количества превращенного в процессе ферментативной реакции белка.
Зависимость оптической плотности от числа единиц активности протеолитических ферментов.
Относя количество введенных в реакцию единиц активности фермента к действию 1 г препарата, определяют его активность. Для расчета прямой, изображенной на графике, составляет расчетное уравнение.
За единицу протеолитической активности принято такое количество фермента, которое катализирует за 30 минут гидролиз 1 г белка в принятых условиях до продуктов, не осаждающихся трихлоруксусной кислотой. 1г составляет 25% от взятого на ферментативную реакцию белка.
Протеолитическая активность характеризуется числом единиц активности фермента, содержащегося в 1 г препарата и твердых полупродуктов или в 100 мл жидких материалов. Метод предназначен для анализа очищенных ферментных препаратов грибного происхождения и всех полупродуктов, получающихся при их производстве.
Определение пепсина по Ансону и Мирскому. Данный метод основан на следующих принципах. Гемоглобин подвергают воздействию пепсина, оставшийся гемоглобин осаждают трихлоруксусной кислотой. Фенольные свойства остатка (тирозин), определяемые фотометрически, используют как меру активности фермента.
Субстратом служит 2%-ный раствор гемоглобина в 0.06 н. растворе соляной кислоты, 5 мл субстрата нагревают до 250С, прибавляют 1 мл раствора фермента, оставляют в течении 10 мин., добавляют 10 мл 0.3 н. раствора трихлоруксусной кислоты, энергично взбалтывают, фильтруют и 5 мл фильтрата отливают в мерную колбу на 25 мл. Сюда же приливают 10 мл 0.5 н. раствора едкого натра и 3 мл фенольного реактива (реактив Фолина-Чиокалтеу разводят предварительно двойным объемом воды) и доводят водой до метки. По истечении нескольких минут окрашенный раствор фотометрируют относительно стандартного раствора, содержащего 0.0008 мэкв тирозина в 5 мл 0.2 н. раствора соляной кислоты (к раствору добавляют 0.5% формальдегида в качестве антисептика). Для исключения возможных ошибок ставят холостой опыт. Единицы пепсина выражают в миллиэквивалентах тирозина (в пределах от 6.10-4 до 11.10-4): единицы пепсина = миллиэквиваленты тирозина х1.47.
Определение протеолитических ферментов по Муру и Штейну. Метод основан на том, что содержащие a-аминогруппу аминокислоты дают с нингидридом окрашенную производную - дикетогидриндилидендикетогидриндамин, а также альдегид-аминокислоты и углекислоту.
Окрашенный продукт обладает характерным максимумом поглощения при 570 мm, а интенсивность окраски зависит от количества аминокислоты. С пролином и оксипиролином реакция протекает в ином направлении, а максимум поглощения получаемого при этом окрашенного продукта расположен при 440 мm.
Инкубацию фермента удобно проводить в мерных колбочках по 5 мл, в которых смешивают и уравновешивают при постоянной температуре на водяной бане все компоненты за исключением фермента. После этого добавляют фермент, раствор смешивают и берут пробы для исследования. Для кинетических измерений концентрацию фермента желательно подобрать таким образом, чтобы аликвотные части можно было бы брать каждые 5 минут.
Аликвотные части пробы, взятые сразу же после добавления фермента, а затем через соответствующие интервалы, добавляют в фотометрические пробирки, содержащие 2 мл реактива нингидрида - это тотчас обрывает реакцию. После этого смесь 20 мин. нагревают на водяной бане; полученная окраска устойчива минимум 24 часа. Средняя ошибка при анализе повторных проб составляет ±2%. После разбавления кипяченой смеси пробы и нингидрида 10 мл смеси, состоящей из равных объемов воды и н.пропанола, интенсивность окраски определяют на спектрофотометре Колемана при 570 мm.
Для калибровки пользуются постоянными количествами стандартных аминокислот, содержащих реакционные компоненты. При помощи калибровочных кривых, построенных отдельно для каждой исследуемой аминокислоты, измерения интенсивности окраски (по отношению к контрольной колбе) пересчитывают в миллимоли. При делении этих величин на миллимоли субстрата, использованного в процессе реакции, можно найти процент гидролиза.
Построение калибровочной кривой исключает ошибки, происходящие от качественных различий реакции аминокислот на цветной реактив. До оптической плотности 0.1 зависимость между величиной поглощения и количеством субстрата носит линейный характер.
Воспроизводимость составляет ±3%. Мешающее влияние аммония, аминов и других веществ, содержащихся в биологическом материале и дающих цветную реакцию с нингидридом, исключается при помощи соответствующих холостых проб. Анализ сравнительно упрощается, когда изучают протеолитические ферменты, субстратами которых являются пептиды, не содержащие свободных аминогрупп. В этом случае вся образующаяся окраска целиком приходится на освободившуюся аминокислоту.
Микрометод определения активности протеиназ А.П.Алексеенко. Предлагаемый микрометод определения активности протеолитических ферментов основан на измерении содержания аргинина в пептидах, освобождающихся при гидролизе протеиназами белковых субстратов. Содержание аргинина определяют с помощью стабилизированной и усовершенствованной реакции Сакагучи (хроматограмму погружают в 0.1% раствор 8-гидроксихинолина в ацетоне, высушивают на воздухе, опрыскивают раствором 0.2 мл Br2 в 100 мл 0.5 М NaOH; аргинин и другие гуанидины дают оранжево-красные пятна, тауромицин и гликоциамин - лишь временную окраску), позволяющей определить содержание аргинина в растворах трихлоруксусной кислоты после осаждения и удаления нерастворимых белков. Зная содержание аргинина в белковом субстрате и определяя его количество в перешедших в раствор пептидах, можно рассчитать процент гидролиза белкового субстрата изучаемыми протеиназами. В этом состоит главное преимущество метода, поскольку метод Ансона не дает возможности рассчитать процент гидролиза белка, атакуемого протеолитическими ферментами. Из всех существующих методов только метод Мура и Штейна позволяет определить процент гидролиза белковых субстратов, но он имеет ограниченное применение, поскольку требует предварительного проведения полного кислотного гидролиза как белка-субстрата, так и образовавшихся пептидов. В предложенной методике калибровочная кривая строится по растворам свободного аргинина. Введение в реакцию пептидов и белков в виде их биуретовых комплексов повысило чувствительность реакции Сакагучи с остатками аргинина и сделало ее такой же чувствительной, как и для свободного аргинина. В качестве белкового субстрата используют классический субстрат - гемоглобин и окисленный по Сангеру лизоцим. Этот низкомолекулярный белок, содержащий 12.7% аргинина прекрасно расщепляется пепсином (рвется около 30 связей).
Прежде чем приступить к определению активности ферментов в изучаемых тканевых субстратах, необходимо:
1. Определить зависимость активности от рН и выбрать оптимальную реакцию среды для проявления активности изучаемого фермента.
2. Построить график зависимости активности фермента от времени его инкубации с субстратом. выбрать для работы те сроки инкубации, при которых сохраняется линейная зависимость между величиной активности фермента и временем его инкубации.
3. Определить зависимость величины активности от концентрации белка в пробе. Выбрать такие концентрации фермента, при которых величина активности фермента была бы пропорциональна его концентрации. Следует учитывать, что в тканевых экстрактах и биологических жидкостях могут присутствовать ингибиторы протеолитических ферментов. При разведении пробы их концентрация снижается , а протеолитических - увеличивается. Определяя “фактор разведения”, можно одновременно с подбором оптимальных условий для определения протеолитической активности выявить и наличие ингибитора.
4. При определении активности фермента необходимо работать при постоянной скорости ферментативной реакции, достигаемой при полном насыщении фермента субстратом, при так называемой максимальной скорости ферментативной реакции. В каждом отдельном случае максимальную скорость необходимо найти экспериментально, измерив активность препарата при разных концентрациях субстрата, при постоянных концентрациях белка и времени инкубирования.
Определение активности протеиназ по белковому субстрату.
К 0.5 мл субстрата в
соответствующем буферном растворе добавляют 0.5 мл пробы, содержащей фермент
(экстракт, биологическую жидкость), инкубируют установленное опытным путем
время , после чего белки в пробе осаждают 5 мл 10% трихлоруксусной кислотой.
Отделяют осадок центрифугированием, а надосадочную жидкость (ТХУ-центрифугат)
подвергают дальнейшей обработке. Контрольные пробы - пробы, где реактив
курсовые - 700 р.
Работы, похожие на Курсовая работа: Определение активности ферментов
|