Реферат: Рекомбинантные вакцины (Генная инженерия)
ОГЛАВЛЕНИЕ
Введение………………………………………………………………………………………….3
1. Принципы конструирования рекомбинантных противовирусных вакцин…………..4-24
1.1. Получение соответствующего фрагмента нуклеиновой кислоты……………....….4
1.2. Выбор высокоактивной и хорошо изученной в иммунологическом отно-
шении модели вектора-носителя и клонирование соответствующего гена……9-21
1.2.1. Получение рекомбинантных ДНК……………………………………………9
1.2.2. Получение рекомбинантных РНК…………………………………………...11
1.2.3. Стратегия клонирования генов………………………………………….…13
1.2.4. Разнообразие векторных молекул…………………………………………17
1.3. Выбор системы экспрессии клонированного гена, способной обеспечить
максимальный выход и функциональную полноценность продукта……………..22
1.4. Создание достаточно удобных и по возможности универсальных векторов
для целевой доставки генов в клетки и ткани организма…………………………23
2. Вакцина против лейкемии кошек, изготовленная с помощью генной
инженерии……………………………………………………………………………….…24
Список литературы…………………………………………………………………………….26
ВВЕДЕНИЕ
Существующие традиционные вакцины, несмотря на очевидный положительный эффект их широкого применения, обладают рядом недостатков. К ним относятся: наличие нежелательных биологически активных и балластных компонентов в препаратах, неполноценные иммунологические свойства самих антигенов. Кроме того, существуют заболевания, не вызывающие иммунитета, вакцины против которых вообще отсутствуют и не могут быть сконструированы на основе классических принципов. Все это вызывает необходимость усовершенствования уже существующих вакцин и создания принципиально новых типов вакцин. Одним из наиболее перспективных направлений в данной области является получение вакцинных препаратов на основе методов генной инженерии.
Последним достижением генной инженерии и биотехнологии стало создание рекомбинантных противовирусных вакцин, содержащих гибридные молекулы нуклеиновых кислот. Данные вакцины обладают целым рядом преимуществ. Они характеризуются отсутствием (или значительным снижением) балластных компонентов, полной безвредностью, низкой стоимостью, которая связана с удешевлением промышленного производства вакцин. Экспрессируемый в клетках вакцинированного животного белок имеет конформацию, близкую к нативной, и обладает высокой антигенной активностью.
Таким образом, рекомбинантные противовирусные вакцины являются новейшим поколением вакцин. Их очевидное преимущество обуславливает широкое применение данного типа вакцин в медицине и ветеринарии для вакцинации населения и сельскохозяйственных животных.
1. ПРИНЦИПЫ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНЫХ ПРОТИВОВИРУСНЫХ ВАКЦИН
Важным условием получения эффективного вакцинного препарата является соблюдение основных принципов его производства. Конструирование рекомбинантных противовирусных вакцин предполагает: 1) получение соответствующего фрагмента нуклеиновой кислоты; 2) выбор высокоактивной и хорошо изученной в иммунологическом отношении модели вектора-носителя и клонирование соответствующего гена (или генов); 3) выбор системы экспрессии клонированного гена, способной обеспечить максимальный выход и функциональную полноценность продукта; 4) создание достаточно удобных и по возможности универсальных векторов для целевой доставки генов в клетки и ткани организма.
1.1. ПОЛУЧЕНИЕ СООТВЕТСТВУЮЩЕГО ФРАГМЕНТА
НУКЛЕИНОВОЙ КИСЛОТЫ
Для конструирования рекомбинантных РНК или ДНК необходимо получить фрагмент нуклеиновой кислоты, который в дальнейшем и будет встраиваться в векторную молекулу.
Фрагменты ДНК для встраивания в вектор можно получить непосредственно из хромосомной ДНК, расщепив ее рестриктазами или разрушив случайным образом (например, с помощью ультразвука) на сегменты с примерно одинаковой длиной. Выделение генов с помощью "вырезания" из генома, как правило, состоит из четырех этапов: 1) получение клонотеки фрагментов генома; 2) выявление фрагментов генома, содержащих необходимый ген, и точная локализация гена в данном фрагменте; 3) вырезание гена из фрагмента(ов) с помощью рестриктаз и сшивка участков гена с помощью ДНК-лигазы фага Т4, если эти участки получены из различных фрагментов; 4) амплификация гена в составе векторной молекулы. Указанный способ получения генов является наиболее приемлимым применительно к протозойным возбудителям, бактериям и для некоторых сложно устроенных ДНК-содержащих вирусов. Такие операции проводятся, в частности, при создании так называемых "библиотек генов", то есть набора одинаковых векторных систем, в совокупности несущих в себе весь геном данного организма. Однако этот подход имеет ряд существенных недостатков. Во-первых, очень сложна задача подбора рестриктаз, позволяющих вырезать из геномной ДНК или клонированного фрагмента генома цельный ген. Как правило, вместе с геном остаются фланкирующие его лишние нуклеотидные последовательности, что мешает дальнейшему использованию этого гена, или же рестриктазы отрезают часть гена, делая его функционально неполноценным. Во-вторых, создание клонотеки генома возбудителя представляет специальную задачу и требует больших затрат времени. Наконец, для ряда ДНК-содержащих вирусов (паповавирусы) доказан сплайсированный характер структуры генов. Вполне понятно, что выделенные гены этих вирусов вследствие наличия интронных областей не будут проявлять функциональной активности в бактериальных клетках и окажутся непригодными при решении задач по конструированию рекомбинантных противовирусных вакцин. В-третьих, если ген составляет незначительную долю от всей геномной ДНК, то возникают большие трудности с его изоляцией и идентификацией.
Наиболее распространенным является путь получения генов через синтез ДНК-копий (кДНК) информационных или каких-либо других (в оптимальном случае - индивидуальных) РНК путем их обратной транскрипции. Данный способ включает в себя три этапа: 1) выделение высокоочищенных мРНК, кодирующих индивидуальные структурные белки, либо выделение геномных РНК вирусов; 2) синтез двухцепочечной ДНК (гена) на матрицах РНК; 3) амплификация гена с помощью методов молекулярного клонирования.
Как отмечалось, первым этапом в получении генов с помощью методов обратной транскрипции служит выделение и очистка геномных РНК и мРНК. Геномные РНК выделяют главным образом из очищенных вирионов, а мРНК - из инфицированных клеток. Для выделения мРНК из инфицированных клеток используют различные способы. В одном из вариантов выделение мРНК из инфицированных клеток проводят непосредственно из лизированных клеток (лизис осуществляют в денатурирующих средах в целях предотвращения разрушающего действия нуклеаз на мРНК) с последующей экстракцией фенолом и хлороформом или центрифигурированием лизата через подушку 5,7M СsCl (для освобождения от клеточных ДНК) и заключительной аффинной хроматографией на олиго(dТ)-целлюлозе (поли(У)-сефарозе). Выделение мРНК можно проводить и из очищенных полирибосом инфицированных клеток.
В случае необходимости получения специфических мРНК, кодирующих индивидуальные структурные белки возбудителя, используют следующий способ. Инфицированные вирусом клетки разрушают механическим путем или химическими методами (использование неионных детергентов). Клеточный лизат освобождают от ядер и митохондрий методом дифференциального центрифугирования и подвергают хроматографии на антительном сорбенте. При этой процедуре с антителами, специфичными к определенному структурному белку возбудителя, связываются полисомы, осуществляющие синтез этого белка. Также может быть использован метод непрямой иммунопреципитации полисом, при котором комплекс антитело - полисома преципитируется из раствора добавлением второго антитела, специфичного к первому. В качестве второго антитела чаще всего берут фиксированные формалином клетки Staphilococcus aureus, на поверхности которых находится так называемый белок А, имеющий сродство к Fc-фрагментам Ig. Из полисом, полученных одним из описанных способов, далее уже выделяют мРНК.
Другой путь выделения индивидуальных мРНК базируется на свойстве мРНК взаимодействовать с комплементарными ДНК, связанными с твердым носителем (целлюлоза, сефароза, нитроцеллюлозные фильтры). Этот метод отбора и очистки специфических мРНК является самым эффективным. Однако его применение возможно только при наличии соответствующих комплементарных ДНК.
Получение препарата очищенной мРНК позволяет перейти к работам по синтезу гена с помощью обратной транскрипции, которую осуществляют с помощью ревертазы AMV в специально подобранных условиях. В процессе обратной транскрипции матрицей служит мРНК, а затравкой олиго(dT12-18) (для мРНК, имеющих поли(А)-тракт) или химически синтезированный олигонуклеотид, комплементарный 3'-концу мРНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (чаще используется обработка щелочью) осуществляют синтез второй цепи ДНК. В этой реакции матрицей служит первая цепь ДНК. Реакция может катализироваться как ревертазой, так и ДНК-полимеразой.
После получения двухцепочечной кДНК следует стадия получения гена, заключающаяся в гидролизе одноцепочечного участка ДНК, соединяющего первую и вторую цепи, нуклеазой S1.
Для получения необходимых участков РНК используют две группы способов: 1) расщепление полирибонуклеотидной цепи РНК в заданном месте; 2) синтез РНК (ферментативный на ДНК- или РНК-матрицах и химический).
Расщепление полирибонуклеотидной цепи РНК может осуществляться в присутствии различных ферментов. Для этой цели используют ферменты: рибонуклеазу H, нуклеазы, ковалентно связанные с олигонуклеотидами (например, стафилококковая нуклеаза), рибозимы (например, рибозим L-19).
Исторически первым способом направленной ферментативной фрагментации РНК (называемой также адресованной, сайт-направленной или сайт-специфической фрагментацией РНК) является разработанный в Московском университете метод гидролиза РНК ферментом РНКазой H в присутствии комплементарных олигодезоксирибонуклеотидов. Этот метод до сих пор остается наиболее универсальным способом направленной фрагментации РНК.
Метод основан на свойстве РНКазы Н расщеплять полирибонуклеотидную цепь РНК в составе ДНК-РНК-гетеродуплекса. К участку РНК, по которому планируется провести ее фрагментацию, синтезируется комплементарный олигодезоксирибонуклеотид длиной в 6-10 нуклеотидных остатков. Далее получают комплекс этого олигонуклеотида с РНК, который затем обрабатывают ферментом. В работе обычно используют РНКазу Н из Escherichia coli - вполне доступный фермент, который может быть довольно легко очищен от всех сопутствующих нуклеазных примесей. Этот метод нашел достаточно широкое применение для сайт-специфического расщепления вирусных и рибосомных РНК, а также для идентификации продуктов процессинга некоторых РНК.
Важным достоинством рассматриваемого метода является и то, что в результате гидролиза РНК рибонуклеазой Н образуются фрагменты, у одного из которых на 5'-конце содержится фосфатная группа, а у другого 3'-конец свободен (нефосфорилирован). Такие фрагменты можно прямо использовать в реакции ферментативного лигирования.
Серьезным ограничением этого метода является то, что участок РНК, с которым связывается комплементарный ему олигодезоксирибонуклеотид, должен иметь однотяжевую конформацию и находиться на поверхности макромолекулы РНК, представляющей собой в условиях расщепления компактную глобулу с развитой вторичной структурой. В отдельных случаях это ограничение удается преодолеть, используя достаточно длинные олигодезоксирибонуклеотиды (15-20-членные), которые предварительно отжигают с частично или полностью денатурированной РНК.
Другое ограничение метода фрагментации полирибонуклеотидов РНКазой Н в присутствии комплементарных олигодезоксирибонуклеотидов заключается в том, что в общем случае предсказать, какая из фосфодиэфирных связей в гетеродуплексе (или в непосредственной близости от него) подвергнется расщеплению, не удается. Более того, фермент зачастую гидролизует не одну, а несколько соседних межнуклеотидных связей. Ясно, что для последующего конструирования рекомбинантных РНК такие фрагменты могут оказаться непригодными.
Расщепление РНК может проводиться и с участием нуклеаз, ковалентно связанных с олигонуклеотидами. Идея, лежащая в основе данного подхода к направленной фрагментации РНК, восходит к концу 60-х годов, когда Н. И. Гриневой и ее сотрудниками был предложен метод олигонуклеотид-направляемой модификации нуклеиновых кислот. Принцип этого метода заключается в том, что с 5'- или 3'-концевым остатком олигонуклеотида, комплементарного заданному району ДНК и РНК, связывают модифицирующий агент, который после образования дуплекса атакует одно из ближайших к нему оснований. Реагентами этого типа удалось направленно фрагментировать фенилаланиновую тРНК из дрожжей, РНК-компонент (M1 РНК) РНКазы Р, а также 16S рибосомную РНК E.coli.
Однако, как и в случае РНКазы Н, расщепление РНК олигонуклеотид-нуклеазой зачастую проходит по нескольким межнуклеотидным связям, что несколько ограничивает возможности применения этого метода для получения рекомбинантных РНК.
Для расщепления РНК применяют также рибозимы (природные РНК и синтетические полирибонуклеотиды, способные катализировать целый ряд превращений у других РНК). Первый рибозим, напоминающий по своим свойствам эндонуклеазы рестрикции, был получен Т. Чеком. В научной литературе его обозначают как рибозим L-19. Этот рибозим представляет собой РНК длиной в 395 нуклеотидных остатков, в 5'-концевой области которой имеется гексануклеотидная последовательность GGAGGG, ответственная за специфичность расщепления предшественника 26S РНК при самосплайсинге. Эта последовательность комплементарна CUCUCU последовательности, расположенной на 3'-конце первого экзонного участка предшественника 26S РНК. Если эту РНК заменить другой, но обязательно содержащей доступную для комплементарного связывания CUCUCUA последовательность, то рибозим L-19 в присутствии гуанозина или гуаниловых нуклеотидов со свободной 3'-гидроксильной группой расщепит ее.
Источником необходимых участков РНК может служить такой способ, как синтез фрагментов РНК. Ферментативный синтез сегментов РНК осуществляют с использованием разнообразных генно-инженерных конструкций. Однако в настоящее время в подавляющем большинстве случаев для препаративного получения РНК используются РНК-полимеразы, закодированные в геномах ряда ДНК-содержащих бактериофагов (Т3, Т7 и SP6). Они характеризуются очень высокой активностью и, в отличие от клеточных РНК-полимераз, состоят из одной полипептидной цепи. Важно также то, что инициация и терминация синтеза РНК этими полимеразами происходит на одном определенном нуклеотидном остатке.
Полученные одним из способов фрагменты нуклеиновых кислот в дальнейшем встраивают в векторные молекулы.
Подводя итог изложенному, можно сделать вывод, что получение генов протективных белков возбудителей представляет собой достаточно сложную задачу. Однако ее решение необходимо для создания в конечном счете рекомбинантных противовирусных вакцинных препаратов.
1.2. ВЫБОР ВЫСОКОАКТИВНОЙ И ХОРОШО ИЗУЧЕННОЙ В
ИММУНОЛОГИЧЕСКОМ ОТНОШЕНИИ МОДЕЛИ ВЕКТОРА-НОСИТЕЛЯ И КЛОНИРОВАНИЕ СООТВЕТСТВУЮЩЕГО ГЕНА
1.2.1. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ ДНК
Суть конструирования рекомбинантных ДНК заключается во встраивании фрагментов ДНК, среди которых находится интересующий нас участок ДНК, в так называемые векторные молекулы ДНК (или просто векторы) - плазмидные или вирусные ДНК, которые могут быть перенесены в клетки про- или эукариот и там автономно репли-цироваться. На следующем этапе проводится отбор тех клеток, которые несут в себе рекомбинантные ДНК (с помощью маркерных признаков, которыми обладает сам вектор), и затем индивидуальных клонов с интересующим нас сегментом ДНК (используя признаки или пробы, специфичные для данного гена или участка ДНК).
При решении ряда научных и биотехнологических задач конструирование рекомбинантных ДНК требует также создания систем, в которых обеспечивается максимальная экспрессия клонируемого гена.
Существует три основных способа встраивания чужеродной ДНК в векторные молекулы. В первом случае 3'-концы фрагментов ДНК, среди которых находится интересующий нас участок ДНК (ген или его сегмент, регуляторный район), с помощью фермента терминальной нуклеотидилтрансферазы наращиваются гомополинуклеотидной последовательностью (например, поли (Т)). 3'-концы линейной формы векторной ДНК тем же способом наращиваются комплементарной ей гомополинуклеотидной последовательностью (то есть поли (А)). Это позволяет соединить две молекулы ДНК путем комплементарного спаривания искусственно полученных "липких" концов.
Во втором случае "липкие" концы создаются с помощью расщепления молекул ДНК (как векторной, так и содержащей интересующий нас фрагмент) одной из эндонуклеаз рестрикции (рестриктаз). Рестриктазы характеризуются исключительно высокой специфичностью. Они "узнают" в ДНК последовательность из нескольких нуклеотидных остатков и расщепляют в них строго определенные межнуклеотидные связи. Поэтому даже в ДНК больших размеров рестриктазы вносят ограниченное число разрывов.
Третий способ представляет собой комбинацию двух первых, когда липкие концы ДНК, образованные рестриктазой, удлиняются синтетическими последовательностями (рис. 1).
Концы фрагментов ДНК можно превратить в "липкие", наращивая их двутяжевыми олигонуклеотидами ("линкерами"), в состав которых входит участок узнавания рестрикта-
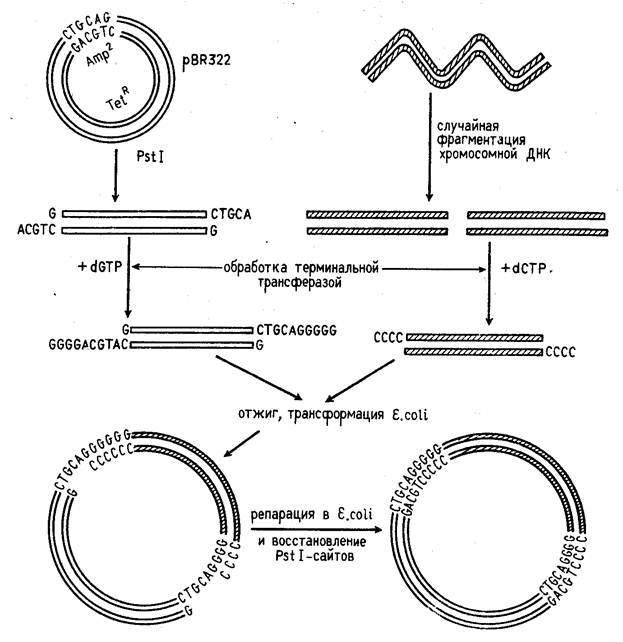
Рисунок 1. Схема конструирования рекомбинантной ДНК с помощью рестриктаз PstI и поли(G)- поли(С)-линкера.
зой. Обработка такого фрагмента данной рестриктазой делает его пригодным для встраивания в векторную молекулу ДНК, расщепленную той же рестриктаэой. Часто в качестве "линкера" применяются полинуклеотидные фрагменты, которые содержат специфические участки сразу для нескольких рестриктаз (их называют "полилинкерами").
После встраивания чужеродной ДНК в вектор их ковалентное сшивание осуществляется ДНК-лигазой. Если же размер бреши в рекомбинированной молекуле превышает одну фосфодиэфирную связь, она застраивается in vitro с помощью ДНК-полимеразы или in vivo с помощью репарирующих систем клетки.
1.2.2. ПОЛУЧЕНИЕ РЕКОМБИНАНТНЫХ РНК
Получение рекомбинантных РНК обычно осуществляют методами ферментативного или химического лигирования РНК. Кроме того, недавно появилась принципиально новая возможность встраивания сегмента РНК в заданное положение других молекул РНК с помощью рибозимов.
Ковалентное сшивание отдельных сегментов РНК при получении рекомбинантных молекул, как правило, осуществляют с помощью Т4 РНК-лигазы. Т4 РНК-лигаза закодирована в геноме бактериофага Т4. Ее выделяют из клеток E.coli, зараженных этим фагом. Фермент сшивает друг с другом однотяжевые олиго- и полирибонуклеотиды. Для работы Т4 РНК-лигазы необходим источник энергии - аденозинтрифосфат. На рис. 2 приведена схема ферментативного лигирования двух коротких олигонуклеотидов. Как видно из этой схемы, акцептором в реакции лигирования служит полностью дефосфорилированный, а донором - полностью фосфорилированный по концевым нуклеотидным остаткам олигонуклеотид. Это предотвращает возможность сшивания однотипных олигонуклеотидов.
Эффективность ферментативного лигирования достаточно длинных полирибонуклеотидов сильно варьирует и ее трудно предсказать исходя только из нуклеотидной последовательности сегментов РНК. Наилучшие результаты получены в тех случаях, когда сшиваемые концы полирибонуклеотидов были пространственно сближены за счет комплементарного связывания соседних с ними участков РНК.
Недавно было установлено, что протяженные сегменты РНК (длиной в 200-300 остатков) могут быть с высоким выходом сшиты Т4 ДНК-лигазой. При этом "стыковка" сегментов осуществляется с помощью олигодезоксирибонуклеотида, комплементарного 3'-концу одного сегмента и 5'-концу другого.
Метод химического лигирования основан на активации концевой фосфатной группы одного из двух сшиваемых сегментов РНК водорастворимым карбодиимидом или
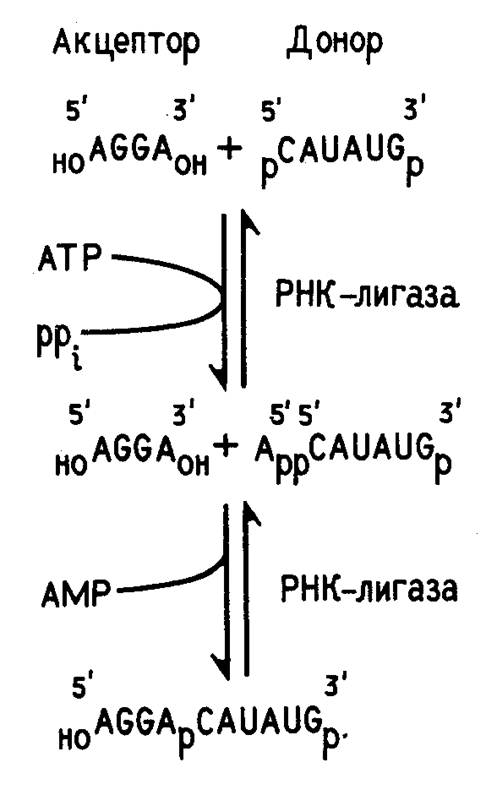
Рисунок 2. Схема сшивания двух олигорибонуклеотидов с помощью Т4 РНК-лигазы.
BrCN. В случае BrCN реакция протекает очень быстро и не сопровождается модификацией нуклеотидных остатков, хотя под действием карбодиимидов фосфодиэфирная связь образуется с более высоким выходом. Для того, чтобы обеспечить сближенность сшиваемых концевых нуклеотидных остатков в фрагментах РНК, было предложено использовать олигодезоксирибонуклеотиды, комплементарные обоим фрагментам в месте их стыка.
Химическое лигирование РНК, как правило, проходит с существенно меньшим выходом, чем ферментативное. Однако оно позволяет получать рекомбинантные РНК с необычными типами межнуклеотидной связи (например, пирофосфатной) и необычными нуклеотидными остатками в месте стыка двух фрагментов.
Получение рекомбинантных РНК с помощью рибозимов основано на обратимости реакции самосплайсинга (при отсутствии гуанозина или гуаниловых нуклеотидов). Это предоставляет возможность для встраивания интронной РНК в заданный участок другого сегмента РНК (рис. 3). Фрагмент РНК, в который производится встраивание, должен содержать нуклеотидную последовательность, идентичную нуклеотидной последовательности 3'-концевого участка 5'-экзонного района 26S РНК и соответственно комплементарную той нуклеотидной последовательности в интроне, которая отвечает за специфичность прямой реакции. Фрагмент, в который производится встраивание, берется в избытке.
В настоящее время описанная здесь цепь реакций может быть реализована только для интронной РНК, получаемой из предшественника 26S РНК тетрахимены. Однако можно думать, что конструирование новых рибозимов может существенно расширить возможности этого подхода.
1.2.3. СТРАТЕГИЯ КЛОНИРОВАНИЯ ГЕНОВ
Векторные молекулы в обязательном порядке содержат маркерные гены, которые после переноса вектора в клетки-реципиенты сообщают им новые свойства. Это может быть устойчивость к антибиотику, которой до трансформации клетки не обладали, или образование фермента, синтез которого в клетках-реципиентах не происходил. Благодаря таким вновь приобретенным признакам клетки с векторными ДНК могут быть легко найдены в популяции исходных клеток. Одновременно могут быть отобраны те клетки, которые содержат векторы со встроенными в них чужеродными ДНК (рекомбинантные ДНК). Для этого встраивание чужеродной ДНК в вектор производится таким образом, чтобы один из маркерных признаков вектора нарушался. Так, например, если бактериальный вектор несет устойчивость к двум антибиотикам, то чужеродную ДНК встраивают в один
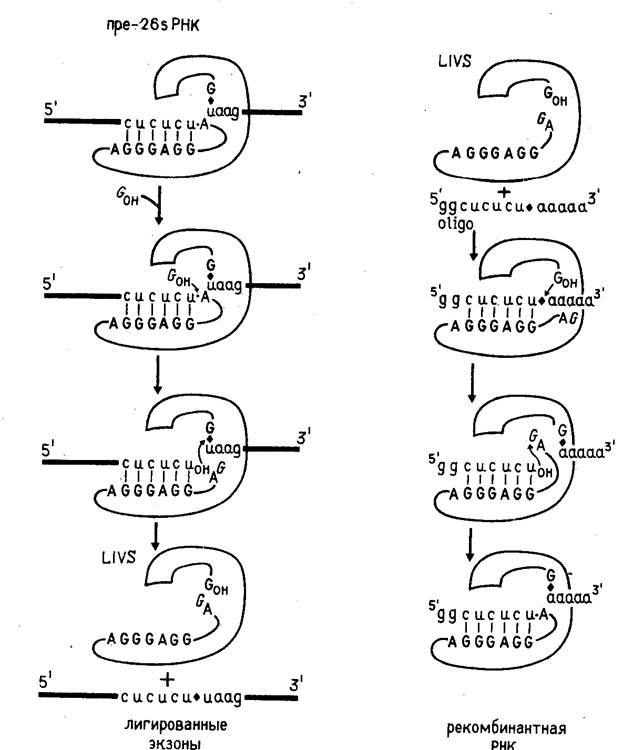
Рисунок 3. Схема прямого и обращенного процесса самосплайсинга.
из генов антибиотической устойчивости. И тогда бактерии с рекомбинантной ДНК, в отличие от бактерий с исходным вектором, могут расти в присутствии только одного из антибиотиков.
Другой весьма распространенный пример связан с наличием в векторной ДНК наряду с генами, сообщающими клетке устойчивость к антибиотикам, фрагмента лактозного оперона, обеспечивающего образование в клетках-реципиентах активного фермента b-галактозидазы. Колонии клеток с таким признаком легко обнаруживаются при выращивании их на твердом агаре, содержащем в качестве субстрата b-галактозидазы 5-бром-4-хлор-3-индолил-b-галактозид (X-gal), поскольку его расщепление приводит к образованию бромхлориндола - красителя, окрашенного в голубой цвет. Если же в ген b-галактозидазы этого вектора встроена чужеродная ДНК таким образом, что этот ген оказался нарушенным, то трансформированные им клетки будут образовывать бесцветные колонии. Само же присутствие рекомбинантного вектора в клетках может быть зафиксировано по их устойчивости к антибиотику.
На следующем этапе среди популяции клеток с рекомбинантными векторами необходимо отобрать индивидуальные клоны, содержащие только интересующие нас гены или их фрагменты. Само собой разумеется, что это в принципе возможно только в том случае, если в исходные клетки проникло в среднем по одной молекуле рекомбинантной ДНК. Способ же отбора клонов в значительной степени зависит от природы клонируемого гена.
По-видимому, самым простым является случай, когда клонируемый ген способен комплементировать ауксотрофную мутацию в штамме-реципиенте. В этом случае клетки высеваются на среду, лишенную вещества, необходимого для роста данного штамма, и только клетки, содержащие рекомбинантную ДНК с искомым геном, способны расти на этой среде. Из таких клонов получают гомогенную культуру клеток, которую используют для получения искомого сегмента ДНК, проделывая все операции в обратном порядке (то есть из клеток выделяют вектор, из него вычленяют необходимый фрагмент ДНК и так далее).
Гораздо чаще для отбора необходимых клонов приходится прибегать к методу ДНК-ДНК- или ДНК-РНК-гибридизации. Для этого необходимо располагать "зондами" (индивидуальными молекулами ДНК или РНК или их фрагментами), комплементарными нуклеотидной последовательности клонируемого гена. Это могут быть специально синтезированные олигодезоксирибонуклеотиды длиной в 15-20 остатков, последовательность которых выбрана на основании полностью или частично известной первичной структуры гена или закодированного в нем белка. Это могут быть кДНК, синтезированные на индивидуальных РНК-копиях данного гена (как таковые или в виде отклонированных, то есть существенно умноженных в количестве фрагментов ДНК). Наконец, это могут быть сами индивидуальные РНК, закодированные в данном гене. Ясно, что во всех случаях "зонды" должны нести радиоактивную метку (обычно 32Р) с достаточно высокой удельной активностью.
Если же индивидуальный "зонд" недоступен, то применяют методы, которые из большого числа рекомбинантов (106) позволяют выбрать сравнительно небольшую группу (около 102), включающую рекомбинант с искомым геном. Эта группа подразделяется на подгруппы (например, на 10) по 10 рекомбинантов. Из каждой подгруппы выделяется ДНК, которую используют для синтеза мРНК и последующей трансляции ее с целью обнаружения соответствующего продукта искомого гена.
Трансляция мРНК может быть осуществлена в бесклеточной системе. Однако в случае эукариотических генов мРНК часто переносят в ооциты лягушки (ксенопуса) с помощью техники микроинъекции, где она транслируется. Продукт трансляции обычно обнаруживают с помощью антител.
Далее в подгруппе, где обнаружен искомый ген, тем же методом исследуется каждый клон.
При наличии зонда с чашки Петри, на которой выращены колонии клеток, делается отпечаток (реплика) на нитроцеллюлозном фильтре. Клеткам дают вырасти на фильтре, затем их разрушают, подвергают ДНК щелочной денатурации и фильтр прогревают при 80°С, после чего ДНК необратимо с ним связывается. Фильтр отмывают от примесей и обрабатывают радиоактивным "зондом" в условиях, оптимальных для ДНК-ДНК- или ДНК-РНК-гибридизации. После удаления избытка "зонда" методом ауторадиографии определяют положение клеточного клона, содержащего участок ДНК с нуклеотидной последовательностью, комплементарной "зонду". Этот клон становится затем источником клеток для получения искомого гена или его фрагмента.
Для селекции клонов, несущих необходимый ген, достаточно широко применяются и иммунологические методы. Принцип отбора на первых этапах тот же, что и при использовании ДНК- и РНК-зондов. Далее с колоний с рекомбинантными ДНК делается реплика с помощью полимерной пластинки, на которой закреплены антитела к продукту искомого гена. Положение клонов, вырабатывающих этот белок, определяется также с помощью антител, но уже меченных радиоактивным йодом (125I).
1.2.4. РАЗНООБРАЗИЕ ВЕКТОРНЫХ МОЛЕКУЛ
Под понятием "вектор" понимается молекула нуклеиновой кислоты, способная после введения в клетку к автономному существованию за счет наличия в ней сигналов репликации и транскрипции.
Векторные молекулы должны обладать следующими свойствами:
1) способностью автономно реплицироваться в клстке-реципиенте, то есть быть самостоятельным репликоном;
2) содержать один или несколько маркерных генов, благодаря экспрессии которых у клетки-реципиента появляются новые признаки, позволяющие отличить трансформированные клетки от исходных;
3) содержать по одному или, самое большее, по два участка (сайта) для различных рестриктаз в разных районах (в том числе в составе маркерных генов), но не в области, ответственной за их репликацию.
В зависимости от целей эксперимента векторы можно условно разделить на две группы: 1) используемые для клонирования и амплификации нужного гена; 2) специализированные, применяемые для экспрессии встроенных чужеродных генов. Вторая группа векторов объединяет векторы, призванные обеспечить синтез белковых продуктов клонированных генов. Векторы для экспрессии содержат последовательности ДНК, которые необходимы для транскрипции клонированных копий генов и трансляции их мРНК в штаммах клеток.
В качестве прокариотических векторов используются плазмиды, бактериофаги; в качестве эукариотических векторов применяют вирусы животных и растений, векторы на основе 2 мкм дрожжей и митохондрий и ряд искусственно сконструированных векторов, способных реплицироваться как в бактериальных, так и в эукариотических клетках (челночные векторы).
Плазмиды - это внехромосомные генетические элементы про- и эукариот, которые автономно реплицируются в клетках. Большинство плазмидных векторов получено на основе природных плазмид ColE1, pMB1 и p15A.
Бактериальные плазмиды делят на два класса. Одни плазмиды (например, хорошо изученный фактор F, определяющий пол у E.coli) сами способны переходить из клетки в клетку, другие такой способностью не обладают. По ряду причин, и прежде всего для предотвращения неконтролируемого распространения потенциально опасного генетического материала, подавляющее большинство бактериальных плазмидных векторов создано на базе плазмид второго класса. Многие природные плазмиды уже содержат гены, определяющие устойчивость клеток к антибиотикам (продукты этих генов - ферменты, модифицирующие или расщепляющие антибиотические вещества). Кроме того, в эти плазмиды при конструировании векторов вводятся дополнительные гены, определяющие устойчивость к другим антибиотикам.
На рис. 4 показан один из самых распространенных плазмидных векторов E.coli - pBR322. Он сконструирован на базе детально изученной плазмиды E.coli - колициногенного фактора ColE1 - и содержит ориджин репликации этой плазмиды. Особенность плазмиды ColE1 (и pBR322 соответственно) состоит в том, что в присутствии ингибитора синтеза белка антибиотика хлорамфеникола (опосредованно ингибирующего репликацию хозяйской хромосомы) ее число в E.coli возрастает от 20-50 до 1000 молекул на клетку, что позволяет получать большие количества клонируемого гена. При конструировании вектора pBR322 из исходных плазмид был делегирован целый ряд "лишних" сайтов для рестриктаз.
В настоящее время наряду с множеством удобных векторных систем для E.coli сконструированы плазмидные векторы для ряда других грамотрицательных бактерий (в том числе таких промышленно важных, как Pseudomonas, Rhizobium и Azotobacter), грамположительных бактерий (Bacillus), низших грибов (дрожжи) и растений.
Плазмидные векторы удобны для клонирования относительно небольших фрагментов (до 10 тыс. пар оснований) геномов небольших размеров. Если же требуется получить клонотеку (или библиотеку) генов высших растений и животных, общая длина генома которых достигает огромных размеров, то обычные плазмидные векторы для этих целей непригодны. Проблему создания библиотек генов для высших эукариот удалось решить с использованием в качестве клонирующих векторов производных бактериофага l.
Среди фаговых векторов наиболее удобные системы были созданы на базе геномов бактериофагов l и М13 E.coli. ДНК этих фагов содержит протяженные области, которые можно делегировать или заменить на чужеродную ДНК, не затрагивая их способности реплицироваться в клетках E.coli. При конструировании семейства векторов на базе ДНК l фага из нее сначала (путем делений коротких участков ДНК) были удалены многие сайты рестрикции из области, не существенной для репликации ДНК, и оставлены такие сайты в области, предназначенной для встраивания чужеродной ДНК. В эту же область часто встраивают маркерные гены, позволяющие отличить рекомбинантную ДНК от исходного вектора. Такие векторы широко используются для получения "библиотек генов". Размеры замещаемого фрагмента фаговой ДНК и соответственно встраиваемого участка чужеродной ДНК ограничены 15-17 тыс. нуклеотидных остатков, так как рекомбинантный фаго -

Рисунок 4. Детальная рестрикционная карта плазмиды pBR322.
вый геном, который на 10% больше или на 75% меньше генома дикого l фага, уже не может быть упакован в фаговые частицы.
Таких ограничений теоретически не существует для векторов, сконструированных на базе нитчатого бактериофага М13. Описаны случаи, когда в геном этого фага была встроена чужеродная ДНК длиной около 40 тыс. нуклеотидных остатков. Известно, однако, что фаг М13 становится нестабильным, когда длина чужеродной ДНК превышает 5 тыс. нуклеотидных остатков. Фактически же векторы, полученные из ДНК фага М13, используются главным образом для секвенирования и мутагенеза генов, и размеры встраиваемых в них фрагментов намного меньше.
Эти векторы конструируются из реплекативной (двутяжевой) формы ДНК фага М13, в которую встроены "полилинкерные" участки (пример такой конструкции показан на рис. 5). В фаговую частицу ДНК включается в виде однотяжевой молекулы. Таким образом, этот вектор позволяет получать клонированный ген или его фрагмент как в двутяжевой, так и в однотяжевой форме. Однотяжевые формы рекомбинантных ДНК широко используются в настоящее время при определении нуклеотидной последовательности ДНК методом Сэнгера и для олигодезоксинуклеотид-направленного мутагенеза генов.
Перенос чужеродных генов в клетки животных осуществляется с помощью векторов, полученных из ДНК ряда хорошо изученных вирусов животных - SV40, некоторых аденовирусов, вируса папиломы быка, вируса оспы и так далее. Конструирование этих векторов проводится по стандартной схеме: удаление "лишних" сайтов для рестриктаз, введение маркерных генов в области ДНК, не существенные для ее репликации (например, гена тимидин-киназы (tk) из HSV (вируса герпеса)), введение регуляторных районов, повышающих уровень экспрессии генов.
Удобными оказались так называемые "челночные векторы", способные реплицироваться как в клетках животных, так и в клетках бактерий. Их получают, сшивая друг с другом большие сегменты векторов животных и бактерий (например, SV40 и pBR322) так, чтобы районы, ответственные за репликацию ДНК, остались незатронутыми. Это позволяет проводить основные операции по конструированию вектора в бактериальной клетке (что технически намного проще), а затем полученную рекомбинантную ДНК использовать для клонирования генов в животной клетке.
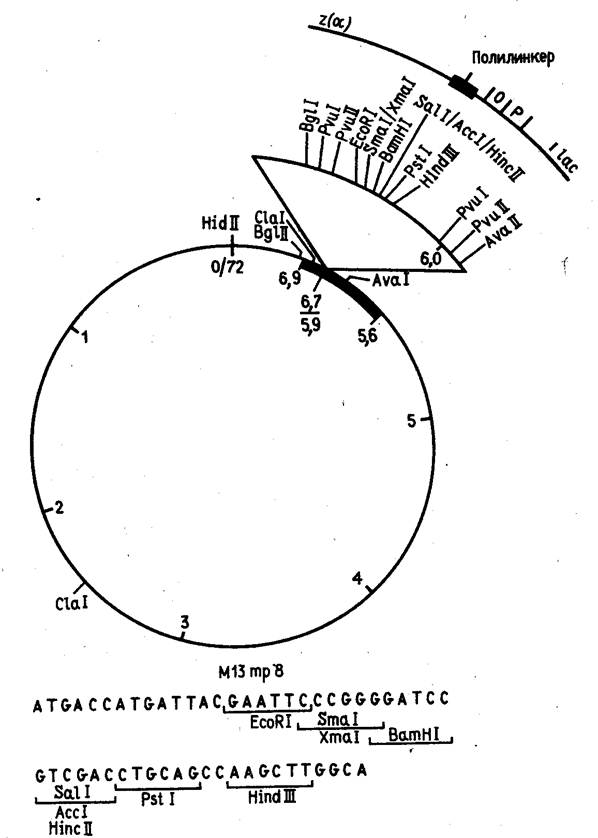
Рисунок 5. Рестрикционная карта вектора М13 mp8.
1.3. ВЫБОР СИСТЕМЫ ЭКСПРЕССИИ КЛОНИРОВАННОГО
ГЕНА, СПОСОБНОЙ ОБЕСПЕЧИТЬ МАКСИМАЛЬНЫЙ ВЫХОД И
ФУНКЦИОНАЛЬНУЮ ПОЛНОЦЕННОСТЬ ПРОДУКТА
Полученные рекомбинантные молекулы переносятся в определенные группы клеток, которые должны обеспечить экспрессию этих генов, то есть синтез соответствующих белков в количествах, экономически рентабельных по сравнению с обычной технологией их производства.
Обычно для данной цели используют бактериальные или дрожжевые культуры клеток, а также системы экспрессии на основе эукариотических клеток.
Из бактериальных клеток наиболее изученной в молекулярно-генетическом отношении является грамотрицательная бактерия Escherichia coli, поэтому для нее можно с наибольшей определенностью планировать генноинженерные конструкции. Однако E. coli слабо освоена промышленностью. Кроме того, она относится к условно-патогенным для человека микроорганизмам, что может создать трудности при получении на ее основе фармацевтических препаратов.
Отмеченные недостатки E. coli легко преодолеваются при конструировании методами генной инженерии штаммов-продуцентов на основе клеток Bacillus subtilis. Данная почвенная бактерия безопасна для человека и животных и прекрасно освоена микробиологической промышленностью. Бактерия B. subtilis по степени изученности следует за E. coli. Важное отличие ее от E. coli - способность эффективно секретировать во внешнюю среду целый ряд белков, поэтому особенно интересны работы по созданию штаммов-продуцентов B. subtilis, секретирующих чужеродные белки из клеток. Однако данная бактерия имеет свои недостатки: рекомбинантные плазмиды в B. subtilis характеризуются нестабильностью, выражающейся в перестройках и делециях ДНК; бациллы секретируют в культуральную среду большое количество протеаз, что существенно усложняет вопрос максимализации генноинженерного получения целевого белка на основе бацилл-продуцентов.
Среди эукариотических микроорганизмов наиболее изученным является низший эукариот Saccharomyces cerevisiae. Одно из преимуществ S. cerevisiae как экспериментальной системы - простота и надежность ее генетического анализа. В клетках дрожжей имеется ферментативная система гликозилирования белков, которая обеспечивает возможность синтеза в них полноценных белков высших эукариот. Аналогичных систем процессинга белков в бактериальных клетках нет. Многие штаммы дрожжей освоены микробиологической промышленностью. Доказана их безвредность для человека и животных. Именно на S. cerevisiae создан первый штамм-продуцент поверхностного антигена вируса гепатита Б, позволивший получить и испытать вакцину против данного вирусного заболевания человека.
С появлением генной инженерии внимание многих исследователей привлекла система культивируемых клеток животных. Особый интерес к культурам клеток животных стал проявляться после обнаружения того, что часть эукариотических генов раздроблена и лишь в системе клеток высших эукариот можно достичь правильной экспрессии таких генов. Кроме того, многие белки животных и их вирусов синтезируются первоначально в виде более высокомолекулярных предшественников, которые в результате специфического протеолитического процессинга переходят в так называемую зрелую форму. Такой процессинг данных белков, по-видимому, можно ожидать лишь в системе клеток животных. Все это убедительно доказывает важность разработки экспрессирующей системы на основе клеток животных.
Для обеспечения наиболее эффективной экспрессии клонированных генов в векторные молекулы встраивают определенные фрагменты ДНК, позволяющие увеличить выход чужеродного белка. Так, для достижения более высокого уровня экспрессии гена HBsAg в клетках E. coli были использованы различные по силе промоторы (промоторы генов cat, kan, bla, trp и тандемно расположенных промоторов генов kan и trp). Уровни синтеза последовательностей HВsAg (нативного и в составе химерных белков) составляли в зависимости от используемых конструкций векторов от 100 до 100000 молекул на клетку.
1.4. СОЗДАНИЕ ДОСТАТОЧНО УДОБНЫХ И ПО ВОЗМОЖНОСТИ
УНИВЕРСАЛЬНЫХ ВЕКТОРОВ ДЛЯ ЦЕЛЕВОЙ ДОСТАВКИ ГЕНОВ В
КЛЕТКИ И ТКАНИ ОРГАНИЗМА
Важным моментом при конструировании ДНК-вакцин является проблема целенаправленной доставки генов в необходимые клетки и защиты вводимых ДНК от действия нуклеаз крови. В результате экспериментальной работы были созданы разнообразные конструкции, позволяющие доставлять целевые гены в клетки-мишени.
Одной из подобных конструкций является модель молекулярного вектора для доставки генов в такие клетки, как лимфоциты и кераноциты. В качестве модельного был использован ген, кодирующий гибридный белок: фактор некроза опухолей-альфа - интерферон-гамма. В центре вектора находится интактная плазмидная ДНК, содержащая доставляемый ген, а на поверхности располагаются антитела к клеткам-мишеням. Конъюгат полиглюкина со спермидином и антителами применяется для связи компонентов (положительно заряженный спермидин обеспечивает связывание конъюгата с плазмидной ДНК). Описанный молекулярный вектор позволяет целенаправленно доставлять гены в клетки-мишени, сводя до минимума их попадание в другие виды клеток, защищать доставляемые гены от нуклеаз крови и использовать положительно заряженный комплекс спермидин-полиглюкин в качестве стимулятора проникновения ДНК в клетки.
В настоящее время также создана векторная модель для доставки в клетки костного мозга гена, кодирующего гранулоцитарный колониестимулирующий фактор человека (чГ-КСФ). Данный белок относится к семейству гемопоэтических факторов роста и является одним из физиологических регуляторов, специфически и высокоэффективно стимулирующих пролиферацию и дифференцировку гемопоэтических предшественников нейтрофилов. чГ-КСФ увеличивает продолжительность жизни клеток костного мозга, усиливает функциональную активность зрелых нейтрофилов. Созданный вектор представляет собой многослойную конструкцию. "Центральным ядром" конструкции является плазмида pGGF8, содержащая ген чГ-КСФ. Ее окружает полисахаридная оболочка, которая состоит из полиглюкина и спермидина. Внешний белковый слой содержит смесь сывороточного альбумина и белка доставки - трансферина. Эффективность описанной векторной модели была доказана опытным путем.
Итак, при конструировании рекомбинантных противовирусных вакцин немаловажное значение имеет создание специального вектора-носителя, обеспечивающего адресную доставку генов и их защиту от действия нуклеаз крови.
2. ВАКЦИНА ПРОТИВ ЛЕЙКЕМИИ КОШЕК,
ИЗГОТОВЛЕННАЯ С ПОМОЩЬЮ ГЕННОЙ ИНЖЕНЕРИИ
Возбудителем лейкемии кошек является ретровирус типа С. Вирус лейкемии кошек (FeLV) имеет в качестве генетического материала молекулу РНК. Заболеваемость и смертность среди домашних кошек в Швейцарии довольно высока. Вакцина против FeLV, изготовленная традиционным путем, была разработана несколько лет тому назад и находится в продаже в Швейцарии. Высокая цена современных вакцин и отсутствие уверенности в отношении длительности эффекта и надежности вакцины вызвали необходимость создания вакцины с помощью генной инженерии.
При конструировании рекомбинантной вакцины оболочечный протеин вируса (env-ген) клонировался в пивных дрожжах (Saccharomyces cerevisiae). Env-ген вначале лигировался с промотором дрожжей (пируваткиназный промотор), а затем клонировался в так называемом "Shuttle"-векторе. Вектор "Shuttle" (челночный вектор) может размножаться как в бактерии E. coli, так и в пивных дрожжах S. cerevisiae.Он содержит с одной стороны репликационные оригины как для дрожжей, так и для E. coli, а с другой стороны селекционный маркер для дрожжей (leu2 ген) и для E.coli (резистентность к ампицилину). Из литра дрожжевой культуры выделяют множество миллиграмм env-протеина и затем тестируют в опыте по вакцинации. Результат вселяет надежду: 10 кошек были иммунизированы env-протеином, причем 4 дали ответ антителами. Через 2 недели провели заражение FeLV. Теперь все животные давали ответ антителами.
Таким образом, полученная вакцина оказалась эффективным средством для борьбы с данным заболеванием.
СПИСОК ЛИТЕРАТУРЫ
1. Грен Э. Я., Пумпен П. П. Рекомбинантные вирусные капсиды - новое поколение иммуногенных белков и вакцин // Журнал ВХО. - 1988. - т. II, №5. - с. 531-536.
2. Дмитриев Б. А. Проблемы и перспективы создания синтетических вакцин // Иммунология. - 1986. - №1. - с. 24-29.
3. Лебедев Л. Р., Сизов А. А., Масычева В. И., Карпенко Л. И., Рязанкин И. А. Молекулярный вектор для доставки генов в клетки-мишени // Биотехнология. - 2001. - №1. - с. 3-12.
4. Мертвецов Н. П., Беклемишев А. Б., Савич И. М. Современные подходы к конструированию молекулярных вакцин. - Новосибирск: Наука, 1987, 210 с.
5. Сизов А. А., Лебедев Л. Р., Масычева В. И., Кашперова Т. А., Одегов А. М. Разработка вирус-подобной конструкции для рецептор-опосредованного транспорта гена чГ-КСФ в клетки костного мозга in vivo // Биотехнология. - 2001. - №1. - с. 13-18.
6. Шабарова З. А., Богданов А. А., Золотухин А. С. Химические основы генетической инженерии. - М.: Изд-во МГУ, 1994, 224 с.
7. Щелкунов С. Н. Клонирование генов. - Новосибирск: Наука, 1986, 232 с.
8. Юров Г. К., Народицкий Б. С., Юров К. П. Конструирование и использование ДНК-вакцин // Ветеринария. - 1998. - №12. - с. 25-27.
9. Hubscher U. Генная инженерия и ветеринария. Вакцины, изготовленные с помощью генной инженерии, и анализ высоковариабельных участков ДНК. -Schweiz. Arch. Tierheilk., 1987, 129, №11, 553-564.
| Продукты рекомбинации: характеристика и манипулирование | |
|
Продукты рекомбинации: характеристика и манипулирование После отбора клонированных рекомбинантных молекул нужно охарактеризовать содержащуюся в них ... Прежде всего нужно очистить рекомбинантную ДНК от ДНК клеток хозяина, РНК и белков. В настоящее время ряд поступающих в продажу ферментов, используемых для конструирования рекомбинантных молекул ДНК, в свою очередь синтезируются под контролем генов, клонированных ... |
Раздел: Рефераты по биологии Тип: курсовая работа |
| Генная инженерия | |
|
СОДЕРЖАНИЕ Введение. 2 ГЛАВА 1. Теоретические предпосылки формирования генной инженерии как науки. 3 1.1. Открытие двойной структуры ДНК и матричного ... Принцип применения ДНК-вакцин заключается в том, что в организм пациента вводят молекулу ДНК, содержащую гены, кодирующие иммуногенные белки патогенного микроорганизма. По своему действию ДНК-вакцины напоминают живые аттенуированные вирусные вакцины или некоторые рекомбинантные вакцины на основе живых вирусных векторов, так как иммуногенные белки ... |
Раздел: Рефераты по биологии Тип: реферат |
| Возникновение злокачественных опухолей | |
|
Министерство образования Российской Федерации Уральский Государственный Технический Университет - УПИ Кафедра Технологии органического синтеза РЕФЕРАТ ... Такие манипуляции осуществляются с помощью соответствующих ферментов - рестрикционных эндонуклеаз, расщепляющих молекулы ДНК в строго определенных участках, и лигаз, сшивающих ... Со времени выдающегося открытия Уотсона и Крика, то есть с середины 50-х годов нашего века, бесспорной считалась основная догма биологии, согласно которой гены, заключенные в ... |
Раздел: Рефераты по биологии Тип: реферат |
| Перспективы применения липосомальных форм | |
|
Оглавление Введение Глава 1. История открытия липосом Глава 2. Применение липосом в медицине 2.1 Свойства липосомальных частиц 2.2 Использование ... С этой целью используют гуманизированные МКА, для удаления мышиного белка, а также Fab' фрагменты антител для удаления фрагментов Fc. Это особенно желательно для применения в фармацевтической области, в частности для доставки материалов, таких как полимерные или белковые лекарственные средства, ДНК-содержащие ... |
Раздел: Рефераты по медицине Тип: дипломная работа |
| Достижения генной инженерии и биотехнологии | |
|
Государственный университет управления Институт государственного и муниципального управления Специальность государственное и муниципальное управление ... Генная инженерия представляет собой совокупность методов, позволяющих не только получать реконбинантные ДНК из фрагментов геномов разных организмов, но и вводить такие ... Для экспрессии в бактериальной клетке гена из клетки животного необходимо, чтобы в клетку была введена молекула ДНК с последовательностью нуклеотидов, кодирующей белок, из которой ... |
Раздел: Рефераты по науке и технике Тип: реферат |