Реферат: Рост и развитие видов рода Acer в условиях интродукции
Базилевская Н. А. Теории и методы интродукции растений. – Москва, 1964
Булыгин Н. Е. Фенологические наблюдения над древесными растениями. – Ленинград, 1979
Дендрология и лесоведение. – Минск, 1967
Дендрологические исследования в заполярье. – Апатиты, 1987
Жизнь растений т.5. – Москва
Кищенко И. Т. Рост и развитие аборигенных и интродуцированных видов семейства Pinaceae Lindl. в условиях Карелии. – Петрозаводск, 2000
Лантратова А. С. Анализ дендрофлоры Карелии. – Петрозаводск, 1985
Лантратова А. С., Овчинникова Е. А. Определитель деревьев и кустарников Карелии. – Петрозаводск, 1978
Мисник Г. Е. Сроки и характер цветения деревьев и кустарников. – Киев, 1976
Флора СССР т.14. – Москва, 1949
Шнелле Ф. Фенология Растений. – Ленинград, 1961
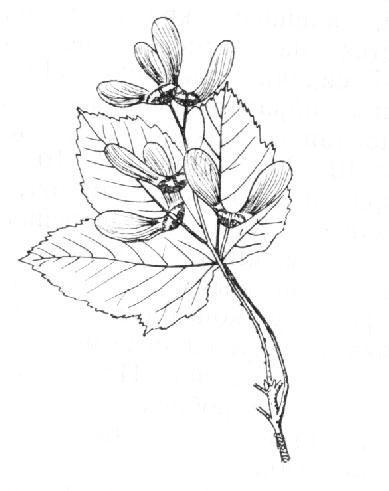
Рис. 2 Acer pseudoplatanus L. – Клен ложноплатановый
Внешний вид побега
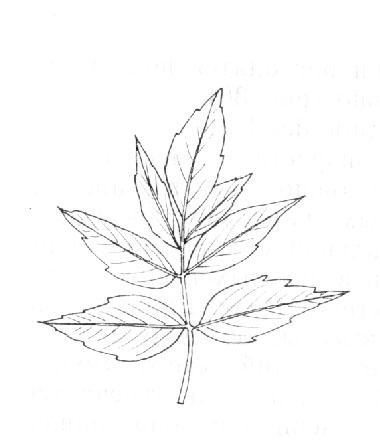
Рис. 3 Acer negundo L. – Клен ясенелистный
Внешний вид побега
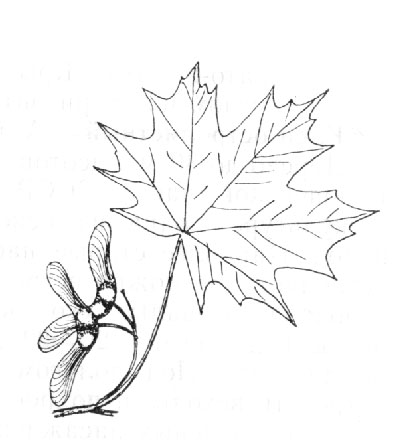
Рис. 1 Acer platanoides L. – Клен остролистный
Внешний вид побега
Введение
Одной из основных задач ботаники является организация мероприятий по охране и обогащению растительного мира. Усиливающееся загрязнение среды, особенно в крупных городах, увеличение размаха строительства дорог и водоемов свидетельствуют о возрастании актуальности озеленительных работ.
Устойчивость к загазованности и задымлению, высокая декоративность, неприхотливость выдвигают многие виды Aceraceae на первое место среди перспективных древесных интродуцентов. Известно, что местные древесные породы часто не могут успешно противостоять ядовитым промышленным выбросам. Они болеют и даже погибают.
Между тем введение в местную флору устойчивых в этом плане растений семейства кленовые поможет аборигенным растениям выдержать этот пресс. К тому же нередко оказывается, что хорошо подобранные к новым экологическим условиям экзоты обладают в 2 – 3 раза большей продуктивностью, чем местные виды.
Хищническое уничтожение растительных ресурсов, загрязнение биосферы отходами производства, ускоренная урбанизация приводят к сокращению ареалов, а иногда и исчезновению самих видов многих растений. В связи с этим, введение некоторых видов растений в культуру является единственным гарантом их сохранения на земле.
Перспективность интродуцентов оценивается по ряду показателей, важнейшими из которых являются сведения об особенностях их фенологического развития.
В Карелии загрязнение окружающей среды постоянно усиливается и небогатая дендрофлора не успевает нивелировать этот процесс, особенно в условиях городов. Поэтому, изучение развития древесных, в том числе и интродуцентов рода Acer для республики весьма актуально. Подобные исследования здесь ранее не проводились.
Глава I Природные условия района исследований
Территория Карелии занимает юго-восточную часть Балтийского кристаллического щита, сложенного докембрийскими горными породами: гранитами, диабазами, кварцо-песчанниками и др. Рельеф данной территории имеет многоскладчатое строение, сложившееся в результате длительных тектонических процессов, происходивших в дочетвертичный период. Для него характерно обилие рек и озер. Ледники, продвигающиеся с северо-запада на юго-восток, сгладили вершины возвышенностей и заполнили впадины продуктами выветривания, вследствие чего образовались моренные гряды, холмы, озы. В целом территория Карелии представляет собой равнину с преобладающими высотами до 100 – 150 м над уровнем моря.
Климат Карелии обуславливается ее территориальным расположением в северных широтах. Солнечная радиация характеризуется сравнительно малой интенсивностью. Благодаря большой продолжительности летнего дня, радиационный баланс деятельной поверхности земли за вегетацию составляет 27.3 ккал/см2.
В южной Карелии положительная среднесуточная температура воздуха наступает в конце апреля, а оттаивание верхнего горизонта почвы – в начале мая. Относительная влажность воздуха в республике довольно высока (75%), причем зимой она в два раза выше, чем летом.
На территории Карелии формируются болотные, подзолистые, болотно-подзолистые почвы. Подстилающими породами являются озерные и моренные пески, суглинки и глина, на которых развиваются подзолистые почвы с железистым и гумусово-железистым иллювиальным горизонтом (Марченко, 1962).
Такое разнообразие почвенного покрова Карелии объясняется климатическими условиями, пестротой форм рельефа, частой сменой почвообразующих пород, характером растительного покрова, степенью заболоченности поверхности.
Как правило, почвы Карелии характеризуются низким плодородием, высокой кислотностью, пониженной температурой в зоне корней (Марченко, 1962).
Природно-климатические условия Ботанического сада, типичны для южной Карелии. Общий климатический режим обусловлен близостью Севера. Солнечная радиация характеризуется малой интенсивностью, однако, благодаря большой продолжительности летнего дня, радиационный баланс деятельной поверхности земли за вегетацию составляет довольно значительную величину (27.3 ккал/см2).
Особенности климата определяются как количеством приходящей солнечной радиации, так и антициклонов. Циклоны сопровождаются потеплением, увеличением количества осадков и повышением влажности воздуха. Антициклоны приносят похолодание.
Весной движущиеся с севера воздушные массы часто приводят к резкому похолоданию, в результате чего затягивается таяние снега и отодвигается начало вегетации растений. В южной Карелии положительная среднесуточная температура воздуха наступает в конце апреля, а оттаивание верхнего горизонта почвы – в начале мая. Однако нередко во второй половине мая температура опускается ниже 0С.
Период со среднесуточной температурой воздуха выше +5С составляет 145 – 155 дней, а с температурой более +10С – 100 – 110 дней. Температура выше +15C отмечается с конца июня до начала третьей декады августа, составляя всего 45 – 50 дней. Суточные колебания температуры воздуха значительно сглаживается в связи с наличием большого количества рек и озер на юге Карелии. Аналогичное влияние на климат, особенно в летнее время, оказывают обширные лесные массивы.
Почвы относятся к подзолистым с железистыми горизонтами на песчаных отложениях, с содержанием большого количества гальки и низким содержанием микро - и макроэлементов.
Для Ботанического сада характерны специфические экологические условия.
Ботанический сад расположен на северном берегу Петрозаводской губы Онежского озера в 6 км северо-восточнее от Петрозаводска, занимая слабый склон южной экспозиции.
Южная экспозиция склона, который занят ботсадом, обеспечивает растениям сравнительно больший приток солнечной радиации и защищает их от холодных северных и восточных ветров.
Благодаря расчлененности рельефа, с большим количеством валунов и скал, на территории сада создается хороший дренаж местности. Близкое залегание грунтовых вод наблюдается лишь в южной части склона, который дополнительно увлажняется поверхностно стекающими водами с вышележащих участков.
Средняя годовая температура –2.4C. Средняя температура наиболее теплого месяца (июля) +16.6C, самого холодного (февраля) –9.9C. Безморозный период обычно начинается со второй половины мая.
Существенное влияние на погодные условия ботсада оказывает близость Онежского озера. Таяние льда в весенний период и аккумуляция тепла холодными водами несколько снижают температуру воздуха на прилегающих прибрежных территориях, тем самым затягивая начало вегетации. Зато осенью, отдавая накопленное за лето тепло, воды озера способствуют повышению влажности воздуха.
Окружающие дендрарий лесные насаждения служат естественной ветрозащитной полосой, предохраняющей посадки от холодных ветров.
Таким образом, особые микроклиматические условия ботсада благоприятствуют работам по интродукции растений, в том числе древесных.
Глава II. Объекты и методика исследований
Род Acer насчитывает приблизительно 120 видов и 40 подвидов, распространенных очень широко в северном полушарии – от субарктических районов в Европе и на Аляске до тропиков Центральной Америки (горы Гватемалы) и Южной Азии. Это листопадные (как исключение, вечнозеленые) деревья. Листья супротивные, простые или иногда сложные. Простые листья бывают лопастными или реже цельными. Сложные листья обычно перистые, с небольшим количеством листочков (3 – 7). Цветки мелкие, собранные преимущественно в кистевидные или метельчатые соцветия, обоеполые, ложнообоеполые (морфологически двуполые, но вследствие недоразвитости тычинок или пестика функционально однополые) и раздельнополые. Растения полигамные. Околоцветник двойной, 4 – 5 членный, иногда редуцирован, у большинства кленовых довольно невзрачный, блеклый, зеленого или желтовато-зеленого цвета. Обычно в цветке клена 5 чашелистиков, 5 лепестков, 8 – 10 тычинок и гинецей из 2 плодолистиков, завязь верхняя (* K0-5 C0-5 A8-10 G(2)). Плод – дробная крылатая двусемянка, при созревании распадающаяся на две односемянные крылатки; семена без эндосперма, с хорошо развитыми семядолями. Большинство видов семейств цветут одновременно с распусканием листьев или позже, энтомофильны, хорошие медоносы. Некоторые виды опыляются ветром, цветут до облиствения. Плоды распространяются ветром.
Все виды кленов являются прекрасными декоративными деревьями и кустарниками, которые ценятся за разнообразие формы листьев и кроны и особенно осенней окраски листьев, принимающей разнообразные оттенки желтого, оранжевого, бронзового и красного цветов. Хорошие медоносы. Древесина кленов обладает всеми качествами хорошей поделочной древесины, плотностью, равномерным строением, прочностью, хорошо полируется, равномерно колется, отличается красивым рисунком и цветом. Поэтому она находит широкое применение в токарном и столярном производствах, а древесина некоторых видов употребляется и на постройки. Некоторые виды (A. velutinum, A. pseudoplatanus) особенно ценятся за хорошие резонаторные свойства древесины, которая и идет на деки для музыкальных инструментов. Дает топливо и уголь высокого качества. Все виды в соке ствола содержат сахар, некоторые, особенно ряд американских видов в таком значительном количестве, что до настоящего времени используются в Канаде для промышленного получения кленового сахара, имеющего своих любителей. Из наших видов сахаром наиболее богаты A. platanoides L., A. pseudoplatanus L. и A. tataricum L. Листья кленов идут на корм скоту и содержат красящие вещества, которые по протравам дают черный цвет, а также коричневые и желтые тона.
Виды рода Acer являются образователями или компонентами широколиственных и смешанных лесов, кустарниковых зарослей. В нашей стране естественно растут клены 27 – 28 видов и более 50 видов интродуцировано.
Acer platanoides L. – Клен остролистный (рис. 1)
Дерево до 20 м, изредка до 30 м высотой с густой, широкой кроной и темной буровато-серой, до почти черной, корой, в зрелом возрасте трескающейся вдоль; листья 5 – 12 (18) см длиной, 8 – 13 (22) см шириной, сверху темно-зеленые, снизу несколько более светлые и блестящие, голые или с нижней стороны с волосками по жилкам и с бородками в углах их, в очертании округлые, обычно 5 – лопастные, реже 3 – или 7 – лопастные; три верхние их лопасти почти равны, нижние значительно мельче их, все крупно выемчато зубчатые, зубцы и верхушки лопастей оттянуты в тонкое остроконечие; основание листьев широко сердцевидное, реже и обычно лишь на коротких побегах усеченное или широко клиновидное. Цветки крупные, зеленовато-желтые, распускающиеся раньше листьев; чашелистики обратно овальные, тупые, лепестки немного уже их, суженные в ноготок, по длине почти равные чашелистикам; лепестки немного уже их, суженные в ноготок, по длине почти равные чашелистикам; крылатка до 4 см длиной, с крыльями, расходящимися под тупым углом, или горизонтально распростертыми.
Распространены в лиственных и смешанных лесах, обычно в качестве примеси, редко в значительном количестве, в горах Кавказа поднимается до субальпийского пояса.
Acer pseudoplatanus L. – Клен ложноплатановый (рис. 2)
Высокое стройное дерево с густой пирамидально-шаровидной кроной, до 40 м высотой и в обхвате до 1.5 м. Кора буровато-серая, у старых деревьев растрескивающаяся и отслаивающаяся, обнажая молодую, светлую кору, отчего стволы кажутся беловатыми или светло-серыми, молодые побеги голые, буровато-серые. Наружные почечные чешуи желто-зеленые, внутренние густо опушенные, желтоватые, крупные. Листья крупные, до 17 см длиной и шириной, округло сердцевидные, сверху темно-зеленые, тусклые, снизу сизоватые или беловатые, иногда грязновато-красные (f. purpurascens Pax), в молодости с нижней стороны, особенно вдоль жилок, довольно густо опушенные, взрослые – лишь с бородками у основания листа в углах жилок. Изредка встречается форма с листьями снизу сплошь волосистыми (var. Villosum Parl.). Листовая пластинка обычно 5 – лопастная, рассеченная на 1/3 – 1/2, реже глубже. Верхние боковые лопасти почти равные средней, направленные косо вверх и обычно потом отклоненные наружу, выемки между ними острые; нижние лопасти небольшие, иногда сильно редуцированные (f. subtrilobum Schwer.) или вовсе не развиваются. Лопасти яйцевидные или яйцевидно треугольные, коротко или реже длинно заостренные, по краю крупно тупо зубчатые, частью двояко-зубчатые, средняя в верхней части иногда несет по одному крупному лопастевидному зубцу с каждой стороны; черешки приблизительно равны пластинке. Соцветие – длинностебельчатая узкая кисть, в нижней своей части иногда сложная, многоцветковая, до 16 см длиной, с волосистыми осью и цветоножками, превышающими почти в два раза цветок. Прицветники мелкие, опадающие. Цветки до 8 мм в диаметре, желтовато-зеленые, с продолговатыми тупыми чашелистиками и лепестками, почти равными по длине, опушенными внутри и по краю длинно реснитчатыми. Крылатки до 6 см длиной, зрелые – бурые, с округлыми выпуклыми орешками, в молодости довольно густо волосистыми, зрелыми – голыми или с рассеянным опушением.
Дерево преимущественно горных лесов, в Европейской части редко спускается на равнину. Может расти на разных почвах, предпочитая известковую. Хорошего роста достигает на глубоких, умеренно увлажненных почвах. Поселяется и на каменистых местах, на осыпях, в расселинах скал, по берегам рек и ручьев. Встречается обычно одиночными экземплярами, редко образует небольшие насаждения. На Кавказе растет в лесах нижнего и среднего поясов, до высоты 1200 – 1500 м над уровнем моря.
Разводится в парках и садах. Известно свыше 50 форм, огромное большинство которых – садовые, полученные в культуре. Среди них следует отметить f. pyramidale Nich. с пирамидальной кроной, f. erythrocarpum Cass. – с красными крылатками, желтовато и бело-пестролистные формы.
Acer ginnala Maxim. – Клен приречный
Маленькое дерево или высокий кустарник, обычно около 2 м высотой, редко до 6 м, с серой гладкой или слегка продольно морщинистой корой и голыми сизовато – зелеными или красноватыми молодыми побегами, позднее становящимися бурыми. Почечные чашелистики у основания красноватые, по краю и на вершине черные. Лепестки блестящие, сверху, темно-зеленые, снизу несколько более светлые, в молодости часто по жилкам волосистые, потом голые, овальные или продолговато овальные, 3 – 8 см длиной, на стерильных до 14 см длиной, 3 – лопастные, с крупной средней лопастью, к основанию обычно суживающейся и длинно заостренной на верхушке, и горизонтальными или слегка направленными кверху яйцевидными острыми или заостренными боковыми лопастями, расположенными на уровне нижней трети пластинки. Края неровно и остро зубчатые, частью двояко зубчатые. На плодущих побегах нижние лепестки большей частью цельные, на длинных стерильных побегах, наоборот, сильнее расчлененные до почти рассеченных. Основание пластинки округлое или слабо сердцевидное, изредка клиновидное. Черешок короче пластинки. Соцветие – густая, овально щитковидная метелка, содержащая 20 – 60 цветков, с слегка пушистыми осями и цветоножками, усаженными, кроме того, коротко стебельчатыми железками. Цветки около 6 мм в диаметре, желтоватые. Крылатки 2.2 – 3 см длиной, зеленые или ярко-розовые, с крыльями, расходящимися под очень острым углом.
Растет зарослями или отдельными группами по берегам рек и речек на песчано-каменистой почве. Заходит также и в горные долины, где селится лишь на открытых местах, в кустарные же заросли на склонах и в лесах не заходит.
Очень декоративно и довольно часто разводится в садах и парках. Из листьев китайцы добывают черную краску под названием «синца».
Acer semenovii Rgl. – Клен Семенова
Кустарник или небольшое деревцо с серой продольно морщинистой корой и голыми бурыми или коричневыми годовалыми побегами. Листья тонкие, но плотные, тусклые, сизо-зеленые, с более светлой нижней стороной, 1.2 – 4.5 см длиной и 1 – 3.2 см шириной, яйцевидные. На плодущих побегах верхние листья 3 – лопастные, с большой средней долей и гораздо более мелкими, косо вверх направленными, редко горизонтальными боковыми, нижние листья нередко цельные, даже иногда цельнокрайние. Лопасти яйцевидные, постепенно заостренные, средняя в верхней части с парой крупных лопастевидных зубцов. Края просто или частью двояко-зубчатые; зубцы туповатые, реже острые. На стерильных побегах листья до 9 –10 см глубокорассеченные, у основания часто с добавочной парой направленных вниз небольших лопастей и 2 парами крупных зубцов на вершине средней лопасти. Основание листьев почти всегда округлое, на стерильных – сердцевидное или реже усеченное. Соцветие – густая щитковидная метелка с осями и цветоножками, довольно густо усаженными коротко стебельчатыми железками. Цветки желтоватые. Крылатки 2.8 – 3.5 см длиной, с орешками в молодости шерстисто опушенными и усаженными железками, зрелыми – обычно голыми. Их крылья наверху расширенные и закругленные, в молодости ярко-розовые, зрелые светло-желтые, расходящиеся под очень острым углом, часто почти параллельные, с внутренними краями нередко соприкасающимися или частично налегающими друг на друга.
Распространен по горным речным долинам, по ущельям и склонам гор, преимущественно в лесах и кустарниковых зарослях, но также и на открытых луговинах, на высоте до 3000 м над уровнем моря.
Acer tataricum L. – Клен татарский
Дерево или высокий кустарник с гладкой темно-серой или почти черной корой и красноватыми или коричневатыми побегами, в молодости пушистыми. Листья 6 – 10 см длиной, 3 – 7 мм шириной, сверху темно-зеленые, снизу несколько более светлые, по жилкам волосистые, от продолговато яйцевидных (f. oblongofolium Rac.) до почти округлых (f. rotundifolium Rac.). Цельные или слегка трехлопастные, по краю неровно двоякопильчатые, иногда волнистые (f. cuspidatum Pax, садовая). Основание чаще неглубоко сердцевидное, реже усеченное. Черешки волосистые, короче пластинки. Цветки в густых овально – щитковидных метелках с волосистыми и усаженными железками осями и цветоножками, с желтоватыми чашелистиками и белыми лепестками. Крылатки 3 – 4 см длиной с голыми (f. tupicum Bionsk. и f. decalvens Blonsk.) или пушистыми (f. hebecarpum Blonsk. и f. pubigerim Blonsk.) орешками, всегда усаженными железками и к основанию суженными, кверху расширенными крыльями, которые то являются прямостоячими с почти параллельными наружными краями (f. genuinum Rac.).
Распространен в подлеске лесов и по опушкам одиночными деревьями или небольшими группами, в лесах по склонам балок и оврагов, по высоким берегам и в поймах рек, на приречных террасах и по склонам гор в зарослях кустарников.
Acer negundo L. – Клен ясенелистный (рис. 3)
Дерево до 25 м высотой и до 1 м в обхвате, часто образующее несколько стволов, с серой корой и с широкой развесистой, поникающей кроной. Молодые побеги голые, зеленые, с густым сизым налетом, ветви серые. Листья из 3 – 5 листочков на черешках 4 – 8 см длиной, черешки боковых листочков обычно не длиннее 1 см, конечного 1.5 – 2.5 см. Листочки более светлые внизу, при распускании с обеих сторон и особенно густо снизу войлочно опушенные, потом голые, 5 – 13 мм длиной, 2.5 – 7.5 см шириной, яйцевидные до эллиптически ланцетных, обычно с оттянутой в остроконечие верхушкой и с равномерно зубчатыми краями, изредка почти цельнокрайние, конечный листочек нередко трехлопастный. Цветки распускаются ранее листьев, тычиночные сначала в плотных пучках, затем цветоножки сильно удлиняются (до 4 – 7 см длиной) и повисают. Чашечка маленькая 1.5 мм длиной, узко колокольчатая, из 5 до половины сросшихся листочков, как и цветоножка, волосистая. Тычинки с волосовидными нитями и линейными пыльниками 3 – 3.5 мм длиной. Пестичные цветки в редких повислых кистях, с чашелистиками у основания сросшимися. Крылатки до 3 см длиной, расходящиеся под прямым или острым углом, с узкими линейными орешками с выдающимися жилками и обычно с согнутыми, к основанию суженными, на вершине расширенными и закругленными крыльями.
Широко распространен в культуре как в Европейской части, так и на Кавказе и в Средней Азии. Родина – Северная Америка.
Разводится в качестве декоративного растения в садах и парках, употребляется для обсадки улиц и дорог, а также для степных лесных посадок. Отличается быстрым ростом и засухоустойчивостью.
Фенологические наблюдения будут проводиться в течение вегетационных периодов 2001 – 2002 годов на территории Ботанического сада Петрозаводского Государственного Университета, расположенного на северном берегу Петрозаводской губы Онежского озера (подзона средней тайги).
Наблюдения должны проводиться через каждые 2 – 3 суток с момента набухания почек до начала опадания листьев.
Должны регистрироваться даты наступления следующих фенофаз: начало набухания и разверзания вегетативных почек, начало и окончание линейного роста побегов, начало опробковения побегов и обособление листьев на побегах, завершение роста и вызревания листьев, начало расцвечивания и опадания листьев, обособление на побегах почек. Вхождение растений в выше перечисленные фазы развития определяется исходя из характеристик, предложенных Н. Е. Булыгиным (1979).
Набухание вегетативных почек. Характеризует начало перехода побегов от этапа внутрипочечного формирования к внепочечному, а растения в целом – от состояния покоя к вегетации. У кленовых набухание почек характеризуется появлением светлых полосок, уголков или каемок между краями наружных чешуй, от чего почки начинают пестреть.
Разверзание вегетативных почек. Характеризует начало внепочечного роста листьев. У видов рода Acer, признаками наступления фенофазы являются появление из-под почечных чешуй, концов зеленых предлистьев – листоподобных образований, переходных от чешуй к настоящим листьям срединной формации.
Начало и окончание роста побегов. Начало фазы отмечается в день, когда становится возможным прощупать пальцами верхушку растущего побега, скрытую в конусе молодых листьев. Окончание роста побегов устанавливается путем измерения их через 3 – 5 дней от нулевой отметки. Побег считается закончившим рост, если по результатам трех последних измерений до основания верхушечной почки длина его не увеличилась. Точность измерения побегов – 5 мм.
Опробковение основания побегов. Фазу отмечают по образованию в нижней части ростовых побегов коркового слоя.
Опробковение ростовых побегов по всей длине. Начинается тогда, когда зеленая окраска растущих побегов сменилась на окраску зрелых побегов, на всем протяжении покрытых пробковой тканью.
Обособление листьев. Фазу отмечают в момент обособления листьев целиком, а не отдельных листочков листа, т. е. в период, когда на черешке сложного листа обособятся и развернутся все листочки. У ряда видов клена первыми на растущих побегах обосабливаются не настоящие листья срединной фармации, а довольно крупные зеленые предлистья, опадающие в начале или середине лета. Это – переходная фаза от разверзания почек к облиствению. Настоящие же листья развертываются лишь спустя 1 – 2 недели после предлистьев, когда и следует отмечать фазу обособления листьев.
Расцвечивание отмирающих листьев. Фаза является индикатором завершения древесными растениями вегетации и перехода их в состояние «покоя».
Опадение листьев. Начало фазы отмечают по появлению под кронами первых опавших листьев.
Обособление на побегах почек. Фаза отмечается в период, когда в пазухах листьев или на концах побегов текущего года можно обнаружить формирующиеся почки, имеющие вид зеленых бугорков.
Набухание репродуктивных почек. Сроки набухания устанавливают, руководствуясь признаками, указанными ранее для вегетативных почек.
Разверзание репродуктивных почек. Фазу устанавливают по появлению из-под разошедшихся внутренних почечных чешуй верхушек зачаточных соцветий, одиночных цветков. Этой фенофазы нет у некоторых представителей рода Acer (Acer tataricum, Acer ginnala), т. к. их цветочные бутоны обнаруживаются лишь в период внепочечного формирования на концах их побегов.
Бутонизация. Фазу отмечают в период, когда в соцветиях или на побегах наблюдается обособление бутонов, т. е. цветков с закрытым околоцветником.
Начало и окончание цветения. В начале цветения полностью раскрывается венчик. При окончании цветения: лепестки завяли, начали усыхать или венчик полностью опал; чашелистики опали или сохраняются в цветке, но усохли.
Глава III Состояние вопроса по теме исследований
Наибольшее влияние на рост и развитие растений оказывают климатические факторы. Особенности ростовых, органообразовательных и физиологических процессов в растительном организме тесно связано с термическими условиями, влажностью среды, световым режимом, количеством осадков.
Дендрофлора Карелии отличается бедностью. Увеличение видового разнообразия флоры невозможно без интродукции растений. В Карелии интродукция растений ведет свое начало с монастырских посадок на о. Валаам, в Петрозаводском летнем саду и городских общественных парках, созданных в конце XVIII – начале XIX вв.
В настоящее время большая часть лиственных интродуцентов сосредоточена в юго-западной Карелии. Общее число садово-парковых насаждений превышает 60 (Марковская и др., 1997).
Раскрытие эколого-биологических особенностей растений невозможно без изучения их сезонного роста и развития. Изучению сезонного развития растений уделяется большое внимание. При этом под развитием растений понимаются «качественные изменения структуры и функций растения и его отдельных частей-органов, тканей и клеток, возникающие в процессе онтогенеза» (Чайлахан и др., 1982). В. Н. Ворошилов (1960) пришел к выводу, что сезонный ритм растений правомерно рассматривать как приспособительное свойство, возникающее в процессе эволюции и закрепляющееся в генотипе для обеспечения оптимального уровня сопряженности во времени процессов роста и развития с сезонными изменениями целого ряда факторов среды обитания. Поэтому все проявления сезонной ритмики (фитоценотические, географические, онтогенетические и погодичные) являются интегральным выражением сложного процесса взаимодействия между эндогенной ритмикой, определяемой генетически заданной программой, и ее экологическими модификациями (Веретенников, 1969; Лархер, 1978; Булыгин, 1979).
Многочисленные публикации свидетельствуют о том, что особенности сезонного развития древесных растений в определенной мере отражают филогенез вида, экологические и адаптационные возможности растений. Было обнаружено, что на конкретном этапе сезонного развития в растении происходят различные физиологические и морфологические изменения. Наблюдающиеся при этом смены фенофаз идут в определенной последовательности, направляемой системой саморегуляции (Лапин и др., 1979). Естественный отбор закрепляет в генотипе прогрессивные изменения биоритмов. Таким образом, для растительных организмов характерны как консерватизм специфичности биоритмов, так и лабильность приспособительных свойств. Соотношения между ними во многом связаны с видовой принадлежностью (Колесниченко, 1985). Как считает автор, особенности этого соотношения и являются одной из основных причин успеха или неудачи интродукции данного вида.
Авторы многочисленных исследований (Ворошилов, 1960; Фролова, 1979) убедительно показали, что особенности различных видов растений обусловлены их неодинаковой требовательностью к экологическим факторам. Поэтому, определив диапазон толерантности той или иной фенофазы к экологическим факторам, можно судить о степени адаптации вида к условиям местообитания.
При анализе степени влияния того или иного фактора среды на начало или окончание фенофазы совершенно необходимо применение математического аппарата, требующего непрерывных наблюдений за средой и фенологией не менее чем за 20 – 30-летний период времени (Зайцев, 1978).
Влияние температуры на рост и развитие. Каждый вид растений, для активной жизнедеятельности после зимнего покоя, требует определенного уровня теплообеспеченности, который называется термическим порогом вегетации (Шульц, 1981). Величина его генетически обусловлена и является видовым признаком. Изменение термического порога вегетации происходит в ряду поколений и составляет главный результат акклиматизации при естественном расселении вида и искусственным переселении при интродукции.
Значение температуры, при которой начинается вегетационный период, зависит как от биологии вида, так и от места происхождения.
Многочисленные исследования свидетельствуют о том, что температура воздуха, особенно в высоких широтах, является основным фактором, непосредственно влияющим на пусковые механизмы, осуществляющие регуляцию ростовых процессов. Начало линейного роста побегов слабо кореллирует с суммой положительных температур. Как правило, в годы с быстрым переходом среднесуточной от отрицательных величин к положительным (при дружной весне), рост побегов начинается при сравнительно небольшой сумме положительных температур, а в годы с затяжной весной – при наибольшей их сумме. В дальнейшем интенсивность роста побегов также связана с температурой воздуха. Как правило, при значительном повышении температуры отмечается и более интенсивный рост побегов. Наступление кульминации прироста побегов тоже связано с температурой воздуха. Он приходится на период с наиболее высокой температурой воздуха за эти месяцы. После кульминации прироста отмечается слабая связь интенсивности роста побегов с температурой воздуха. В этот период она остается еще сравнительно высокой, а прирост снижается довольно быстро. Прекращение роста побегов в незначительной мере зависит от температурного режима. Еще более ослабевает влияние данного фактора на фазу опробковения основания побегов.
Влияние влажности воздуха на сезонный рост и развитие. Влажность воздуха имеет существенное значение для жизни древесных растений. Как отмечает Э. П. Галенко (1976), процесс фотосинтеза у растений наиболее продуктивно протекает при относительной влажности воздуха 60 – 70% при условии, что и другие жизненные факторы среды находятся в оптимуме.
Влияние атмосферных осадков на сезонное развитие и рост. Осадки служат основным источником накопления запасов почвенной влаги. Исследования, проведенные И. Т. Кищенко, показали, что количество и частота выпадения осадков не отражаются на темпах роста вегетативных органов дерева. Осадки оказывают заметное влияние на сроки прекращения деятельности камбия ствола. При этом характер их влияния на этот процесс различается в связи с условиями произрастания. В условиях избыточного почвенного увлажнения формирование древесины длится тем дольше, чем меньше выпадает осадков в период затухания роста, и, наоборот, обилие осадков приводит к более раннему прекращению роста. Совершенно противоположный эффект в условиях умеренной увлажненности почвы.
Влияние солнечной радиации на сезонный рост и развитие. Солнечный свет является источником энергии для фотосинтеза, эффективность которого во многом определяется интенсивностью солнечной радиации, которая может усиливать или замедлять данный процесс. Продолжительность и интенсивность солнечного сияния оказывает воздействие на развитие растений как через изменение скорости продуцирования ассимилятов, так и через изменение температурного режима.
Вероятно, так же как и температура воздуха, солнечная радиация влияет на сроки наступления начала роста терминальных побегов. По мере роста побегов этот фактор продолжает оказывать влияние на интенсивность деятельности апекальной меристемы, однако, по сравнению с температурой воздуха его значение для роста менее существенно (И. Т. Кищенко, 2000).
Урожайность и качество семян. Перспективность интродуцированных растений оценивается по целому ряду признаков, позволяющих судить о степени их адаптации к новым условиям среды. Наиболее существенными из них являются интенсивность и качество семеношения, характеризующие возможность отбора среди интродуцентов особей с высокой генеративной способностью, хорошо адаптировавшимся к новым условиям. Поэтому в интродукционной работе с древесными растениями вопросам семенного размножения уделяется большое внимание.
Заключение
Усиливающееся загрязнение окружающей среды, особенно в крупных населенных пунктах, свидетельствует о необходимости широкомасштабных озеленительных работ. Среди наиболее перспективных древесных интродуцентов многие виды рода Acer имеют в этом плане ряд преимуществ, обладая повышенной устойчивостью к задымлению и загазованности воздуха, высокой декоративностью и способностью выделять фитонциды.
Бедность дендрофлоры многих регионов нашей страны, в том числе и Карелии, настоятельно требует обогащения новыми видами древесных растений, введением их в культуру.
Одним из важнейших показателей успешности адаптации интродуцентов к новым условиям среды является ритмика ростовых процессов в связи с изменчивостью экологических факторов.
Зная особенности конкретного вида интродуцента, можно объективно оценить перспективность их использования в зеленом строительстве.
Курсовая работа «Рост и развитие видов рода Acer в условиях интродукции»
2001 Клементьев Николай Анатольевич (ПетрГУ, каф. Ботаники и физиологии растений, руководитель – профессор, д б н , Кищенко Иван Тарасович)
В настоящий момент ведется работа по изучению данного рода.
Для дополнительной информации пишите nknet@karelia.ru
Приложение 2
Даты наступления фенофаз у Acer ginnala Maxim.
Год наблюдения |
Пг1 |
Пг2 |
Пб1 |
Пб2 |
О1 |
О2 |
Л1 |
Л2 |
Л3 |
Л4 |
| 1976 | 12.V | 17.V | 8.VI | 1.VII | 10.VIII | 24.V | 10.VI | 14.IX | 24.IX | |
| 1977 | 5.V | 13.V | 6.VI | 13.VII | 28.VI | 29.VII | 17.V | 23.V | 6.VII | 21.IX |
| 1978 | 14.V | 18.V | 4.VI | 25.VII | 4.VII | 10.VIII | 22.V | 31.V | 15.VII | 5.X |
| 1979 | 14.V | 18.V | 30.V | 24.VII | 2.VII | 9.VIII | 22.V | 29.V | 17.VII | 25.IX |
| 1980 | 8.V | 22.V | 3.VI | 26.VII | 5.VII | 12.VIII | 28.V | 2.VI | 20.VII | 23.IX |
| 1981 | 11.V | 20.V | 26.V | 22.VII | 30.VI | 8.VIII | 23.V | 25.V | 16.VII | 25.IX |
| 1982 | 6.V | 10.V | 3.VI | 28.VII | 5.VII | 12.VIII | 21.V | 31.V | 21.VII | 23.IX |
| 1983 | 29.IV | 4.V | 31.V | 20.VII | 28.VI | 5.VIII | 16.V | 28.V | 15.VII | 20.IX |
| 1984 | 5.V | 10.V | 27.V | 16.VII | 2.VII | 2.VIII | 18.V | 24.V | 12.VII | 18.IX |
| 1985 | 8.V | 28.V | 12.VI | 30.VII | 8.VII | 15.VIII | 31.V | 10.VI | 25.VII | 29.IX |
| 1986 | 7.V | 15.V | 30.V | 28.VII | 5.VII | 21.VIII | 23.V | 27.V | 18.VII | 20.IX |
| 1987 | 30.IV | 10.V | 1.VI | 5.VII | 12.VIII | 30.V | 7.VI | 29.VII | 1.X | |
| 1988 | 8.V | 17.V | 26.V | 30.VII | 4.VII | 10.VIII | 29.V | 4.VI | 10.VII | 22.IX |
| 1989 | 21.IV | 1.V | 14.V | 19.VIII | 4.VII | 1.IX | 17.V | 28.V | 1.VII |
|
Год наблюдения |
Л5 |
Ц1 |
Ц2 |
Ц3 |
Ц4 |
Ц5 |
Пл1 |
Пл2 |
Пл3 |
Пл4 |
| 1976 | 12.X | 1.VI | 8.VI | 12.VI | 3.VII | 25.VII | 21.VII | 15.VIII | ||
| 1977 | 5.X | |||||||||
| 1978 | 3.X | 2.VI | 9.VI | 16.VI | 27.VI | 20.VII | 4.VII | 12.VIII | ||
| 1979 | 6.X | 31.V | 4.VI | 11.VI | 20.VI | 18.VII | 2.VII | 10.VIII | ||
| 1980 | 6.X | 3.VI | 6.VI | 12.VI | 18.VI | 10.VII | 6.VII | 29.VII | 1.IX | |
| 1981 | 15.X | 1.VI | 5.VI | 11.VI | 19.VI | 8.VII | 4.VII | 30.VII | 3.IX | |
| 1982 | 15.X | 3.VI | 7.VI | 15.VI | 5.VII | 16.VII | 8.VII | 29.VII | 31.VIII | |
| 1983 | 8.X | 30.V | 4.VI | 11.VI | 27.VI | 8.VII | 4.VII | 30.VII | 25.VIII | |
| 1984 | 5.X | 23.V | 29.V | 6.VI | 17.VI | 6.VII | 30.VI | 27.VII | 22.VIII | |
| 1985 | 17.X | 10.VI | 16.VI | 20.VI | 6.VII | 25.VII | 10.VII | 15.VIII | 8.IX | 20.IX |
| 1986 | 6.X | 30.V | 4.VI | 12.VI | 20.VI | 13.VII | 7.VII | 30.VII | 21.VIII | |
| 1987 | 21.X | 30.IV | 16.V | |||||||
| 1988 | 6.X | |||||||||
| 1989 | 21.IV | 1.V | 3.VI | 9.VI | 26.VI | 24.VI |
Приложение 3
Даты наступления фенофаз у Acer semenovii Rgl.
Год наблюдения |
Пг1 |
Пг2 |
Пб1 |
Пб2 |
О1 |
О2 |
Л1 |
Л2 |
Л3 |
Л4 |
| 1976 | 11.V | 17.V | 7.VI | 17.VII | 25.V | 30.V | 24.VIII | 24.IX | ||
| 1977 | 2.V | 5.V | 31.V | 28.VII | 2.VII | 4.VIII | 16.V | 23.V | 11.VII | 20.IX |
| 1978 | 9.V | 18.V | 2.VI | 1.VIII | 8.VII | 8.VIII | 22.V | 31.V | 20.VII | 2.X |
| 1979 | 10.V | 18.V | 31.V | 30.VII | 12.VII | 9.VIII | 22.V | 29.V | 18.VII | 3.X |
| 1980 | 8.V | 17.V | 6.VI | 28.VII | 10.VII | 11.VIII | 26.V | 1.VI | 21.VII | 25.IX |
| 1981 | 8.V | 20.V | 27.V | 27.VII | 8.VII | 8.VIII | 23.V | 25.V | 16.VII | 28.IX |
| 1982 | 3.V | 8.V | 27.V | 3.VIII | 12.VII | 12.VIII | 15.V | 24.V | 21.VII | 25.IX |
| 1983 | 25.IV | 5.V | 29.V | 22.VII | 6.VII | 7.VIII | 13.V | 24.V | 14.VII | 22.IX |
| 1984 | 3.V | 8.V | 25.V | 18.VII | 3.VII | 3.VIII | 16.V | 23.V | 9.VII | 20.IX |
| 1985 | 7.V | 26.V | 7.VI | 5.VIII | 14.VII | 15.VIII | 29.V | 5.VI | 23.VII | 4.X |
| 1986 | 28.IV | 5.V | 27.V | 28.VII | 10.VII | 16.VIII | 11.V | 24.V | 20.VII | 18.IX |
| 1987 | 28.IV | 8.V |
1.V/15.VII |
30.VII/30.IX |
22.VII | 25.VIII | 3.VI | 7.VI | 17.VII | 2.X |
| 1988 | 5.V | 13.V | 23.V | 8.VIII | 3.VII | 3.VIII | 26.V | 4.VI | 10.VII | 20.IX |
| 1989 | 22.IV | 29.IV | 13.V | 22.VII | 29.VII | 16.V | 28.V | 1.VII |
|
Год наблюдения |
Л5 |
Ц1 |
Ц2 |
Ц3 |
Ц4 |
Ц5 |
Пл1 |
Пл2 |
Пл3 |
Пл4 |
| 1976 | 11.X | 25.V | 28.V | 5.VI | 4.VII | 20.VII | 18.VII | 5.VIII | 20.VIII | |
| 1977 | 4.X | |||||||||
| 1978 | 15.X | 28.V | 6.VI | 14.VI | 27.VI | 20.VII | 10.VII | 10.VIII | ||
| 1979 | 15.X | 27.V | 4.VI | 10.VI | 20.VI | 14.VII | 11.VII | 9.VIII | ||
| 1980 | 3.X | 3.VI | 7.VI | 14.VI | 18.VI | 11.VII | 4.VII | 29.VII | 1.IX | |
| 1981 | 30.IX | 28.V | 2.VI | 9.VI | 17.VI | 10.VII | 4.VII | 3.VIII | 28.VIII | |
| 1982 | 15.X | 27.V | 4.VI | 9.VI | 8.VII | 18.VII | 12.VII | 30.VII | 31.VIII | |
| 1983 | 12.X | 26.V | 2.VI | 8.VI | 29.VI | 9.VII | 3.VII | 3.VIII | 25.VIII | |
| 1984 | 10.X | 21.V | 28.V | 4.VI | 12.VI | 6.VII | 30.VII | 30.VII | 20.VIII | 25.IX |
| 1985 | 5.X | 2.VI | 14.VI | 19.VI | 6.VII | 21.VII | 15.VII | 15.VIII | 2.IX | 1.X |
| 1986 | 13.X | 20.V | 2.VI | 10.VI | 18.VI | 15.VII | 10.VII | 30.VII | 25.VIII | |
| 1987 | 28.X | 28.IV | 4.V | 20.VII | 14.VII | |||||
| 1988 | 10.X | |||||||||
| 1989 | 22.IV | 29.IV | 2.VI | 19.VI | 28.VI | 27.VI | 12.VII | 23.VIII |
Приложение 1
Даты наступления фенофаз у Acer platanoides L.
Год наблюдения |
Пг1 |
Пг2 |
Пб1 |
Пб2 |
О1 |
О2 |
Л1 |
Л2 |
Л3 |
Л4 |
| 1976 | 15.V | 19.V | 8.VI | 27.VII | 29.VI | 25.VII | 23.V | 27.V | 18.VII | 6.IX |
| 1977 | 5.V | 13.V | 26.V | 19.VII | 27.VI | 28.VII | 16.V | 23.V | 23.VI | 2.IX |
| 1978 | 14.V | 18.V | 2.VI | 20.VII | 10.VII | 1.VIII | 22.V | 28.V | 6.VII | 10.IX |
| 1979 | 14.V | 18.V | 29.V | 18.VII | 7.VII | 3.VIII | 22.V | 29.V | 5.VII | 10.IX |
| 1980 | 30.IV | 22.V | 6.VI | 20.VII | 10.VII | 29.VII | 30.V | 5.VI | 7.VII | 8.IX |
| 1981 | 2.V | 20.V | 28.V | 18.VII | 8.VII | 25.VIII | 23.V | 26.V | 6.VII | 15.IX |
| 1982 | 3.V | 16.V | 27.V | 21.VII | 11.VII | 19.VIII | 22.V | 26.V | 12.VII | 18.IX |
| 1983 | 27.IV | 18.V | 28.V | 16.VII | 6.VII | 17.VIII | 20.V | 26.V | 8.VII | 15.IX |
| 1984 | 6.V | 16.V | 27.V | 12.VII | 3.VII | 12.VIII | 18.V | 22.V | 6.VII | 10.IX |
| 1985 | 14.V | 27.V | 14.VI | 20.VII | 10.VII | 20.VIII | 1.VI | 12.VI | 12.VII | 25.IX |
| 1986 | 4.V | 16.V | 28.V | 15.VII | 5.VII | 15.VIII | 20.V | 23.V | 10.VII | 12.IX |
| 1987 | 15.V | 15.V | 23.V | 15.VII | 9.VII | 22.VIII | 20.V | 25.V | 15.VII | 29.IX |
| 1988 | 11.V | 20.V | 21.V | 4.VII | 1.VII | 18.VIII | 27.V | 31.V | 1.VII | 10.IX |
| 1989 | 25.IV | 2.V | 10.V | 12.VI | 14.VI | 15.VIII | 13.V | 23.V | 21.VI | 21.IX |
|
Год наблюдения |
Л5 |
Ц1 |
Ц2 |
Ц3 |
Ц4 |
Ц5 |
Пл1 |
Пл2 |
Пл3 |
Пл4 |
| 1976 | 24.IX | 21.V | 23.V | 25.V | 27.VI | 4.VII | 1.VII | 1.VII | 24.IX | |
| 1977 | 26.IX | |||||||||
| 1978 | 5.X | 14.V | 22.V | 25.V | 28.VI | 4.VII | 2.VII | 15.VII | ||
| 1979 | 4.X | 15.V | 23.V | 27.V | 30.V | 20.VI | 9.VI | 10.VII | 25.IX | 15.X |
| 1980 | 1.X | 26.V | 29.V | 1.VI | 3.VI | 10.VI | 6.VI | 11.VII | 28.VIII | |
| 1981 | 30.IX | 21.V | 23.V | 25.V | 29.V | 9.VI | 3.VI | 27.VI | 26.VIII | |
| 1982 | 5.X | 23.V | 25.V | 30.V | 31.V | 17.VI | 9.VI | 26.VI | 29.VIII | |
| 1983 | 5.X | 16.V | 20.V | 24.V | 27.V | 4.VI | 30.V | 27.VI | 10.VIII | |
| 1984 | 2.X | 15.V | 19.V | 21.V | 23.V | 2.VI | 28.V | 13.VI | 6.VIII | 29.X |
| 1985 | 15.X | 28.V | 30.V | 4.VI | 7.VI | 18.VI | 14.VI | 10.VII | 25.IX | 15.X |
| 1986 | 5.X | 16.V | 19.V | 25.V | 26.V | 9.VI | 3.VI | 26.VII | 20.VIII | |
| 1987 | 21.X | 30.IV | 12.V | 20.V | 4.VI | 14.VI | 10.VI | |||
| 1988 | 10.X | 10.V | 19.V | 27.V | 29.V | 11.VI | 7.VI | 23.VII | 1.IX | |
| 1989 | 21.IV | 30.IV | 6.V | 9.V | 23.V | 15.V | 29.VI | 20.VIII |
Приложение 5
Даты наступления фенофаз у Acer negundo L.
Год наблюдения |
Пг1 |
Пг2 |
Пб1 |
Пб2 |
О1 |
О2 |
Л1 |
Л2 |
Л3 |
Л4 |
| 1976 | 3.V | 15.V | 1.VI | 13.VIII | 22.VI | 12.VIII | 19.V | 29.V | 12.VIII | 6.IX |
| 1977 | 5.V | 10.V | 26.V | 22.VII | 20.VI | 30.VIII | 16.V | 23.V | 15.VII | 10.IX |
| 1978 | 9.V | 14.V | 28.V | 30.VII | 12.VII | 30.VIII | 18.V | 22.V | 22.VII | 10.IX |
| 1979 | 10.V | 16.V | 29.V | 27.VII | 10.VII | 29.VIII | 21.V | 27.V | 18.VII | 10.IX |
| 1980 | 30.IV | 14.V | 6.VI | 28.VII | 17.VII | 28.VIII | 26.V | 30.V | 17.VII | 8.IX |
| 1981 | 2.V | 10.V | 25.V | 25.VII | 18.VII | 26.VIII | 22.V | 26.V | 16.VII | 15.IX |
| 1982 | 30.IV | 3.V | 27.V | 30.VII | 19.VII | 30.VIII | 10.V | 25.V | 21.VII | 17.IX |
| 1983 | 24.IV | 30.IV | 22.V | 22.VII | 18.VII | 20.VIII | 3.V | 22.V | 16.VII | 10.IX |
| 1984 | 25.IV | 3.V | 18.V | 20.VII | 12.VII | 18.VIII | 12.V | 23.V | 10.VII | 8.IX |
| 1985 | 4.V | 16.V | 4.VI | 31.VII | 20.VII | 1.IX | 30.V | 7.VI | 22.VII | 20.IX |
| 1986 | 24.IV | 30.IV | 22.V | 29.VII | 18.VII | 12.VIII | 8.V | 25.V | 18.VII | 8.IX |
| 1987 | 27.IV | 4.V | 20.V | 24.VII | 20.VII | 1.IX | 1.VI | 7.VI | 22.VII | 5.X |
| 1988 | 10.V | 17.V | 22.V | 21.VII | 4.VII | 24.V | 27.V | 6.VII | 15.IX | |
| 1989 | 25.IV | 1.V | 9.V | 3.VII | 22.VII | 10.VIII | 14.V | 19.V | 23.VI | 23.IX |
|
Год наблюдения |
Л5 |
Ц1 |
Ц2 |
Ц3 |
Ц4 |
Ц5 |
Пл1 |
Пл2 |
Пл3 |
Пл4 |
| 1976 | 15.IX | 17.V | 18.V | 20.V | 28.V | 7.VI | 8.VI | 3.VII | 10.VIII | |
| 1977 | 28.IX | 13.V | 16.V | 23.V | 4.VI | 13.VI | 11.VI | 8.VII | ||
| 1978 | 30.IX | 18.V | 22.V | 28.V | 2.VI | 10.VI | 6.VI | 20.VII | 2.X | |
| 1979 | 25.IX | 19.V | 22.V | 26.V | 30.V | 2.VI | 31.V | 18.VII | 28.IX | |
| 1980 | 1.X | 20.V | 26.V | 28.V | 2.VI | 9.VI | 6.VI | 18.VII | 28.VIII | |
| 1981 | 29.IX | 18.V | 20.V | 23.V | 26.V | 30.V | 28.V | 17.VII | 27.VIII | |
| 1982 | 8.X | 8.V | 10.V | 12.V | 18.V | 28.V | 24.V | 25.VII | 29.VIII | |
| 1983 | 4.X | 6.V | 9.V | 12.V | 16.V | 24.V | 20.V | 17.VII | 30.VIII | 25.IX |
| 1984 | 2.X | 5.V | 10.V | 14.V | 16.V | 20.V | 18.V | 4.VII | 28.VIII | 18.IX |
| 1985 | 10.X | 14.V | 27.V | 1.VI | 6.VI | 12.VI | 9.VI | 16.VII | 1.X | 25.IX |
| 1986 | 6.X | 6.V | 15.V | 23.V | 23.V | 26.V | 22.V | 18.VII | 30.VIII | 20.IX |
| 1987 | 25.X | 25.IV | 2.V | 12.V | 18.V | 12.VI | 7.VI | 25.VII | 10.IX | 18.IX |
| 1988 | 10.X | 8.V | 15.V | 21.V | 26.V | 18.VI | 3.VI | 20.VII | 1.IX | 22.IX |
| 1989 | 21.IV | 30.IV | 7.V | 10.V | 20.V | 16.V | 18.VI | 15.IX |
Приложение 4
Даты наступления фенофаз у Acer tataricum L.
Год наблюдения |
Пг1 |
Пг2 |
Пб1 |
Пб2 |
О1 |
О2 |
Л1 |
Л2 |
Л3 |
Л4 |
| 1976 | 9.V | 16.V | 10.VI | 10.VIII | 22.VII | 20.VIII | 29.V | 3.VI | 29.VII | 4.IX |
| 1977 | 2.IV | 5.V | 6.VI | 18.VII | 28.VI | 25.VII | 16.V | 31.V | 11.VII | 19.IX |
| 1978 | 9.V | 8.V | 4.VI | 25.VII | 21.VI | 10.VIII | 22.V | 25.V | 15.VII | 25.IX |
| 1979 | 10.V | 16.V | 29.V | 20.VII | 12.VII | 9.VIII | 22.V | 28.V | 10.VII | 23.IX |
| 1980 | 8.V | 22.V | 6.VI | 19.VII | 14.VII | 12.VIII | 28.V | 4.VI | 11.VII | 23.IX |
| 1981 | 8.V | 20.V | 29.V | 18.VII | 15.VII | 6.VIII | 23.V | 27.V | 12.VII | 28.IX |
| 1982 | 30.IV | 5.V | 28.V | 25.VII | 19.VII | 12.VIII | 22.V | 27.V | 21.VII | 25.X |
| 1983 | 24.IV | 3.V | 29.V | 16.VII | 8.VII | 5.VIII | 18.V | 25.V | 10.VII | 5.X |
| 1984 | 3.V | 8.V | 23.V | 12.VII | 6.VII | 3.VIII | 18.V | 21.V | 8.VII | 30.IX |
| 1985 | 6.V | 16.V | 4.VI | 28.VII | 20.VII | 15.VIII | 30.V | 2.VI | 22.VII | 5.X |
| 1986 | 5.V | 5.V | 28.V | 15.VII | 8.VII | 14.VIII | 16.V | 20.V | 15.VII | 30.IX |
| 1987 | 29.IV | 5.V | 20.V | 24.VI | 21.VII | 22.VIII | 4.VI | 10.VI | 20.VII | 16.X |
| 1988 | 8.V | 16.V | 28.V | 3.VII | 1.VII | 15.VIII | 29.V | 3.VI | 29.VI | |
| 1989 | 23.IV | 30.V | 15.V | 26.VI | 24.VI | 10.VIII | 17.V | 24.V | 21.VI |
|
Год наблюдения |
Л5 |
Ц1 |
Ц2 |
Ц3 |
Ц4 |
Ц5 |
Пл1 |
Пл2 |
Пл3 |
Пл4 |
| 1976 | 30.IX | 20.VI | 25.VI | 1.VII | 8.VII | 25.VII | 28.VII | 20.VIII | ||
| 1977 | 3.X | 11.VI | 13.VI | 17.VI | 30.VI | 18.VII | 15.VII | 1.VIII | 29.IX | |
| 1978 | 15.X | 2.VI | 9.VI | 18.VI | 29.VI | 20.VII | 10.VII | 1.VIII | ||
| 1979 | 12.X | 3.VI | 9.VI | 17.VI | 25.VI | 18.VII | 8.VII | 31.VII | 25.IX | |
| 1980 | 10.X | 5.VI | 9.VI | 14.VI | 18.VI | 21.VII | 7.VII | 31.VII | 28.VIII | |
| 1981 | 15.X | 2.VI | 9.VI | 15.VI | 24.VI | 20.VII | 6.VII | 28.VII | 27.VIII | |
| 1982 | 15.X | 2.VI | 8.VI | 16.VI | 6.VII | 26.VII | 20.VII | 30.VII | 29.VIII | |
| 1983 | 16.X | 30.V | 5.VI | 13.VI | 21.VI | 12.VII | 5.VII | 26.VII | 20.VIII | |
| 1984 | 12.X | 23.V | 30.V | 8.VI | 16.VI | 9.VII | 3.VII | 24.VII | 28.VIII | |
| 1985 | 9.VI | 14.VI | 20.VI | 6.VII | 26.VII | 12.VII | 3.VIII | 26.IX | ||
| 1986 | 16.X | 30.V | 7.VI | 16.VI | 25.VI | 15.VII | 10.VII | 12.VIII | 4.IX | |
| 1987 | 25.X | 30.IV | 10.V | 3.VI | 17.VI | 30.VII | 14.VII | 18.VIII | 20.IX | |
| 1988 | 6.V | 14.V | 25.VI | 30.VI | 5.VII | 2.VII | 29.VII | |||
| 1989 | 21.IV | 28.IV | 20.V | 9.VI | 27.VI | 20.VI | 12.VII | 20.IX |
Условные обозначения фенофаз
Пг1 – набухание почек
Пг2 – раскрывание почек
Пб1 – начало роста побегов
Пб2 – окончание роста побегов
О1 – частичное одревеснение побегов
О2 – побеги одревесневают полностью
Л1 – обособление листьев
Л2 – листья не достигают нормальных размеров
Л3 – конец роста и вызревание листьев
Л4 – расцвечивание листьев
Л5 – опадение листьев
Ц1 – набухание почек
Ц2 – разверзание почек
Ц3 – бутонизация
Ц4 – начало цветения
Ц5 – окончание цветения
Пл1 – завязывание плодов
Пл2 – плоды достигают зрелых размеров
Пл3 – созревание плодов
Пл4 – опадение плодов