Реферат: Клонирование и анализ генов легких цепей иммуноглобулинов стерляди
ОГЛАВЛЕНИЕ
СПИСОК СОКРАЩЕНИЙ..............................................................................................3
ВВЕДЕНИЕ.............................................................................................................................4
ОБЗОР ЛИТЕРАТУРЫ......................................................................................................5
СТРОЕНИЕ ИММУНОГЛОБУЛИНОВ...................................................................................5
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ
ГЕНОВ L-ЦЕПЕЙ ИГ У МЛЕКОПИТАЮЩИХ....................................................................7
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ
ГЕНОВ L-ЦЕПЕЙ ИГ У НИЗШИХ ПОЗВОНОЧНЫХ......................................................11
ЭВОЛЮЦИЯ ГЕНОВ L-ЦЕПЕЙ ИГ.......................................................................................19
МАТЕРИАЛЫ И МЕТОДЫ...........................................................................................24
ЭЛЕКТРОФОРЕЗ ДНК............................................................................................................24
ВЫДЕЛЕНИЕ ДНК ИЗ ГЕЛЯ.................................................................................................24
ПОЛУЧЕНИЕ РАДИОАКТИВНЫХ ЗОНДОВ......................................................................25
ВЫДЕЛЕНИЕ ПЛАЗМИДНОЙ ДНК......................................................................................26
ОЧИСТКА ПЛАЗМИДНОЙ ДНК МЕТОДОМ РАВНОВЕСНОГО
ЦЕНТРИФУГИРОВАНИЯ В ГРАДИЕНТЕ ПЛОТНОСТИ CsCl........................................26
ТРАНСФОРМАЦИЯ ПРОКАРИОТИЧЕСКИХ КЛЕТОК...................................................27
ВЫДЕЛЕНИЕ ЛЕЙКОЦИТОВ ИЗ КРОВИ РЫБ.................................................................28
ВЫДЕЛЕНИЕ СУММАРНОЙ РНК ИЗ ЛЕЙКОЦИТОВ.....................................................28
ВЫДЕЛЕНИЕ ВЫСОКОМОЛЕКУЛЯРНОЙ ДНК
ИЗ ЭУКАРИОТИЧЕСКИХ КЛЕТОК.....................................................................................28
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК......................................................................29
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ...............................................................................33
СУБКЛОНИРОВАНИЕ ДНК...................................................................................................34
СКРИНИНГ БИБЛИОТЕКИ кДНК........................................................................................34
ОПРЕДЕЛЕНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК.............................35
САУЗЕРН БЛОТ ГИБРИДИЗАЦИЯ.....................................................................................36
РЕЗУЛЬТАТЫ.....................................................................................................................38
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК И ЕЕ СКРИНИНГ.....................................38
ОПРЕДЕЛЕНИЕ ПЕРВИЧНОЙ СТРУКТУРЫ......................................................................40
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ..............................................40
АНАЛИЗ ГЕНОМНОЙ ОРГАНИЗАЦИИ..............................................................................41
ОБСУЖДЕНИЕ....................................................................................................................46
ВЫВОДЫ...............................................................................................................................48
СПИСОК ЛИТЕРАТУРЫ................................................................................................49
СПИСОК СОКРАЩЕНИЙ
ИГ иммуноглобулины
пн пар нуклеотидов
тпн тысяч пар нуклеотидов
е.а. единица активности
ИПТГ изопропилтиогалактозид
ТЕМЕД N,N,N,'N'-тетраметил-этилен-диамин
трис трис(гидроксиметил)аминометан
дНTФ дезоксиаденозинтрифосфат
ддНТФ дидезоксинеклеотидтрифосфат
дНТФ дезоксинуклеотидтрифосфат
рATФ рибоаденозинтрифосфат
ДТТ дитиотрейтол
Na2ЭДТА этилендиаминтетраацетат натрия
SDS додецилсульфат натрия
X-Gal 5-бромо-4-хлоро-3-индолил-бета-D-галактозид
ВВЕДЕНИЕ
В основе функционирования гуморального иммунитета у млекопитающих лежит сложный комплекс молекулярно-генетических и физиологических механизмов, обеспечивающих реорганизацию, мутагенез и клональную экспрессию генов иммуноглобулинов. Возникновение этой подсистемы иммунитета связывают с появлением хрящевых рыб.
На ранних этапах эволюции гены ИГ были организованы в виде повторяющихся кластеров V-J-C генных сегментов. При подобной организации разнообразие продуцируемых антител основывалось, прежде всего, на количестве имевшихся в геноме кластеров, каждый из которых кодировал один вариант полипептидных субъединиц ИГ. В ходе дальнейшей эволюции произошел переход от кластерной к сегментарной организации, обеспечивающей дополнительный источник разнообразия за счет комбинативной рекомбинации генных сегментов. Предполагается, что этот переход также имел важное значение для регуляции экспрессии генов и осуществления механизмов клонального отбора антителопродуцирующих клеток в ходе иммунного ответа.
Настоящая работа представляет собой часть проекта, направленного на изучение эволюции механизмов изотипического исключения генов легких цепей ИГ у низших позвоночных. Целями работы являлись изучение структуры и организации генов легких цепей ИГ стерляди (Acipenser ruthenus), представителя филогенетически древней группы костно-хрящевых рыб.
ОБЗОР ЛИТЕРАТУРЫ
СТРОЕНИЕ ИММУНОГЛОБУЛИНОВ
Иммуноглобулины выполняют в организме позвоночных функцию гуморальных антител и антиген-связывающих рецепторов В-лимфоцитов. Особенностью этого класса белков является огромное разнообразие, позволяющее им вступать во взаимодействие с фактически любыми биологическими макромолекулами.
Типичная молекула ИГ состоит из двух идентичных тяжелых (H) и двух идентичных легких (L) полипептидных цепей. H- и L-цепи построены из нескольких доменов, каждый из которых состоит примерно из 110 аминокислотных остатков. У млекопитающих существует пять классов H-цепей: g, m, a, d, и e. Каждая H-цепь построена из одного N-концевого и нескольких (трех или четырех) C-концевых доменов. N-концевые домены различаются в разных молекулах и называются вариабельными (V) доменами. C-концевые домены имеют одинаковую структуру у молекул одного класса и называются константными (С) доменами (рис. 1) (Пол, 1987).
Среди L-цепей млекопитающих различают два типа: лямбда и каппа. Каждая L-цепь состоит из одного вариабельного и одного константного доменов. В индивидуальной молекуле ИГ присутствует только один тип L-цепи (Roitt et al., 1993).
Гигантское разнообразие ИГ обеспечивается прежде всего тем, что V и C области кодируются разными генами (генными сегментами), физически разнесенными в зародышевой ДНК.
В формировании вариабельных доменов H-цепей участвуют три генных сегмента: вариабельный (V), D-сегмент (от англ. diversity- разнообразие) и J-сегмент (от англ. joining- соединяющий). С-области Н-цепей разных классов кодируются отдельными генами (Roitt et al., 1993).
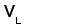
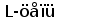

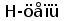




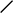
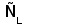

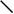
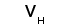



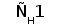

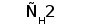
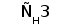
Рис. 1. Схема строения молекулы иммуноглобулина.
Типичная молекула иммуноглобулина содержит две идентичные легкие (L) и две идентичные тяжелые (H) цепи, связанные между собой ковалентно дисульфидными связями. Каждая L-цепь состоит из одного вариабельного (VL) и одного константного (СL) домена. Н-цепь состоит из одного VH и нескольких CН доменов.
В формировании зрелого гена L-цепи участвуют три генных сегмента: V-сегмент и J-сегмент кодируют V-домен, C-сегмент кодирует константный домен. На поздних стадиях развития В-лимфоцита генные сегменты, кодирующие вариабельные домены, объединяются в различных сочетаниях, образуя матрицу для экспрессии L- и H-цепей (Seidman et al., 1979; Durdik et al., 1984).
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ ГЕНОВ L-ЦЕПЕЙ ИГ У МЛЕКОПИТАЮЩИХ
Различные виды млекопитающих имеют сходную схему организации и экспрессии генов ИГ. Одним из наиболее изученных в этом отношении видов является человек. Лямбда и каппа цепи ИГ у человека кодируются генами, расположеными в разных локусах и на разных хромосомах. Каппа локус состоит из большого количества Vk генных сегментов, собраных в группы, пяти Jk и одного Сk сегмента.. Такой тип организации называется сегментарным (рис. 2). Лямбда локус состоит из группы Vl сегментов, точное количество которых неизвестно, и семи пар Jl-Cl сегментов. Три из них (JCl4 - JCl6) являются псевдогенами (Hieter et al., 1981; Roitt et al., 1993).
В процессе развития В-лимфоцита в кроветворных органах один из V сегментов соединяется с одним из J сегментов посредством сайтспецифической рекомбинации (Schatz et al., 1992). В этот процесс вовлекаются специальные олигомерные последовательности: гепта- и нонамеры, фланкирующие с 3' стороны V сегмент и с 5' стороны J сегмент (рис. 3) (Aguilera et al., 1987; Hesse et al., 1989). Отличительной особенностью лямбда и каппа локусов является конфигурация промежутков между гепта- и нонамерами, называемых спейсерами. С V и J генными сегментами лямбда типа ассоциированы 23 пн и 12 пн спейсеры соответственно, а с V и J сегментами каппа типа - 12 пн и 23 пн спейсеры (Durdik et al., 1984; Stavnezer et al., 1985).
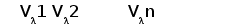
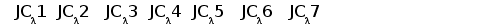

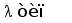



 л
л



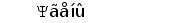


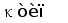

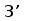
Рис. 2. Схема строения лямбда и каппа локусов генов L-цепей ИГ человека.
Лямбда локус содержит много Vl сегментов и семь пар близкорасположенных Jl-Cl. Три из них яляюся псевдогенами (Y). Каппа локус содержит много Vk пять Jk и один Ck генный сегмент (Hieter et al., 1981; Roitt et al., 1993).
Обозначения:
- сигнальные
олигомеры.
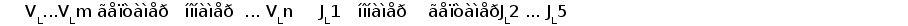
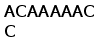


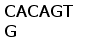


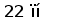
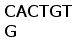
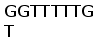
Рис. 3. Схема расположения олигонуклеотидных последовательностей, задействованных в сайтспецифической рекомбинации в каппа локусе млекопитающих. Эти последовательности фланкируют с 3‘ стороны VL сегмент и с 5’ стороны JL сегмент. Во время рекомбинации гептамер и нонамер одного из VL сегмента объединяются с гептамером и нонамером одного из JL сегмента. Это делает возможным соединение VL и JL сегментов без нарушения рамки трансляции (Aguilera et al., 1987; Hesse et al., 1989).
 Обозначения:
- сигнальные
олигомеры.
Обозначения:
- сигнальные
олигомеры.
Последовательность, в которой рекомбинируют локусы L-цепей, строго упорядочена во времени. Показано, что первыми перестраиваются локусы каппа типа, причем если в одной хромосоме перестройка оказалась нефункциональной, тоесть не привела к появлению полноценного гена, то рекомбинация происходит в каппа локусе на гомологичной хромосоме. В случае повторной абортивной перестройки аналогично происходят перестройки в лямбда локусах. Если в клетке непродуктивно перестроились все четыре локуса, то она превращатся в 0-клетку и подвергается апоптозу. Если же перестройка одного из локусов привела к образованию функционального гена, то рекомбинация остальных локусов блокируется. Детали механизма блокирования неизвестны, однако установлено, что необходимым условием для него является транскрипция продуктивно перестроенного гена. В результате в каждом индивидуальном лимфоците синтезируется только один тип L-цепей, каппа или лямбда, и экспрессия гена происходит только на одной из двух гомологичных хромосом. Эти феномены, называемые изотипическим и аллельным исключениями лежат в основе ключевого принципа функционирования иммунной системы - принципа клональной селекции (Пол, 1987; Strob, 1987).
В организме млекопитающих генерируется свыше десяти миллионов вариантов антител, хотя в геноме содержится значительно меньшее количество генных сегментов, которые могут участвовать в формировании вариабельных доменов. В случае L-цепей, основным источником разнообразия является комбинативное сочетание V и J сегментов. Дополнительным источником является смещение рекомбинационной рамки в месте их соединения. Кроме того, V генные сегменты могут обмениваться участками ДНК с псевдогенами посредством генной конверсии. Очень существенный вклад в разнообразие специфичностей антител вносит соматическое мутирование вариабельных генных сегментов в сайтах, участвующих в формировании антигенсвязывающего центра. Соматическое разнообразие генерируется при вторичном иммунном ответе путем включения гипермутационного механизма в клетках памяти в зародышевых центрах лимфатических узлов (Tonegawa 1983; Roitt et al., 1993).
ГЕНОМНАЯ ОРГАНИЗАЦИЯ И ЭКСПРЕССИЯ ГЕНОВ L-ЦЕПЕЙ ИГ У НИЗШИХ ПОЗВОНОЧНЫХ
Хрящевые рыбы.
Гуморальный иммунный ответ в виде специфических антител обнаруживается только у позвоночных. Наиболее примитивными видами, у которых обнаружена способность синтезировать ИГ, являются хрящевые рыбы. Представители трех основных таксонов (акулы, скаты и химеры) продуцируют антитела в ответ на введение широкого спектра антигенов. Однако, в отличие от млекопитающих, гетерогенность сывороточных антител у хрящевых рыб выражена очень слабо. Кроме того, у них не обнаружено созревание иммунного ответа - при вторичном введении антигена спектр антител не меняется (Flajnik, 1996). Результаты исследований последних лет продемонстрировали, что организация генов ИГ у хрящевых рыб также имеет ярко выраженные особенности.
У хрящевых рыб обнаружены гены трех типов L-цепей, локализующиеся в разных локусах. В каждом локусе геные сегменты организованы в кластеры, имеющие по одному V, J и C генному сегменту (рис. 4). Таких кластеров содержится несколько десятков или даже сотен в одном локусе на расстоянии 12-15 тпн друг от друга (Rast et al., 1994).
Гены первого типа найдены у разнозубой акулы (Heterodontus francisci) (Shamblott and Litman, 1989) и малого ската (Raja erinacea) (Anderson et al., 1995). У акулы кластеры имеют размер 3-6 тпн. V и J, J и C генные сегменты разделены примерно 0,5 и 4 тпн соответственно. У ската V и J сегменты слиты в зародышевой ДНК.
Гены второго типа обнаружены у пятнистой химеры (Hydrolagus colliei) (Maisey, 1984), песчаной акулы (Carcharhinus plumbeus) (Hohman et al., 1992; Hohman et al., 1993), разнозубой акулы и малого ската (У всех изученных видов в кластерах этого типа V и J генные сегменты соединены уже в зародышевом геноме. V и C гeнные сегменты разделены 2-3 тпн.
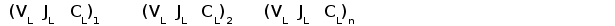





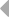
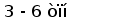




Рис. 4. Схема строения локусов генов L-цепей ИГ у акулы.
Организация генов у акулы имеет кластерный тип. Каждый кластер имеет размер 3 - 6 тпн и содержит по одному VL, JL и CL генному сегменту.
Обнаружено три типа генов L-цепей ИГ. Локусы генов первого и третьего типов имеют схожую организацию. В этом случае VL и JL, JL и CL сегменты разделены примерно 0,5 и 4 тпн соответственно (А) (Rast et al., 1994).
В локусе генов второго типа VL и JL сегменты соединены уже в зародышевом геноме (Б).
Третий тип генов L-цепей найден у акулы-няньки (Gynglimostoma cirratum) (Greenberg et al., 1993) и разнозубой акулы (Rast et al., 1994). В кластерах этого типа V и J, J и C генные сегменты разделены, как и в кластерах первого типа, примерно 0,5 и 4 тпн соответственно. V и J генные сегменты фланкируются сайтами специфической рекомбинации со спейсерами 12 и 23 пн, что соответствует конфигурации спейсеров в локусах каппа типа млекопитающих. По первичной структуре гены третьего типа тоже наиболее близки к генам каппа типа высших позвоночных (гомология по аминокислотной последовательности составляет около 60%).
Гены этих трех типов имеют между собой низкую степень гомологии:40-50% по нуклеотидной последовательности С сегментов и 50-60% по последовательности V сегментов (Hohman et al., 1993).
Полученные данные показывают, что у хрящевых рыб имеется очень большое количество генных сегментов. Однако специфика организации генов исключает комбинативное сочетание сегментов. Все разнообразие антител у хрящевых рыб формируется только за счет экспрессии большого количества кластеров.
Данные о нуклеотидных последовательностях из разных локусов выявили низкую гетерогенность генов (85-95%), что указывает на отсутствие соматического мутагенеза (Rast et al., 1994).
Костистые рыбы.
Анализ сывороточных антител пещерного сомика (Ictalurus punctatus) показал наличие трех типов L-цепей с различными молекулярными массами: 26, 24 и 22 кДа. Два вида антител мыши, 3F12 и 1G7, захватывают более 90% от общего количества иммуноглобулинов в реакции иммунопреципитации. При этом 3F12 антитела связываются с молекулами, содержащими L-цепи массой 24 и 22 кДа, а 1G3 антитела с другой субпопуляцией, содержащей L-цепи массой 26 кДа. На основе этих данных L-цепи разделяют на два класса, называемые F и G (Lobb et al.,1984).
Геномная организация локуса L-цепей G типа имеет кластерный тип (рис. 5). В каждом кластере содержится по одному J и C сегменту и два V сегмена. Вариабельные генные сегменты расположены всегда в противоположной транскрипционной полярности к J и C сегментам . В ряде геномных клонов один V сегмент располагался с 3' стороны от C генного сегмента. Размер кластера равен примерно 3 тпн, расстояние между кластерами около 6 тпн (Ghaffari and Lobb, 1993; Bengten, 1994).
Хотя гены L-цепей сомика имеют низкую гомологию с генами других позвоночных (40-50% по нуклеотидной последовательности), их можно отнести к каппа типу высших позвоночных, так как сходство по первичной структуре с лямбда типом ниже. Кроме того, конфигурация спейсеров имеет характер каппа типа: 12 пн спейсер ассоциирован с V, 23 пн спейсер с J сегментом (Ghaffari and Lobb, 1993).
У радужной форели (Oncorhynchus mykiss), представителя другого семейства костистых рыб, также обнаружены два типа L цепей (Daggfeldt et al., 1993). Один из них (L1) высокогомологичен G типу сомика по структуре V и C областей и соответствующий локус имеет сходную организацию.
По-видимому, у этих видов объединение V и J сегментов происходит только за счет инверсий с восстановлением единой полярности транскрипции. Разнообразие вариабельных доменов образуется в результате комбинативного сочетания V и J сегментов в пределах одного кластера. Не исключается, что в перестройки могут вовлекаться и соседние кластеры.
Интересной особенностью костистых рыб является очень высокая пропорция мРНК, представляющей так называемые стерильные транскрипты (Ghaffari and Lobb, 1993). Эти транскрипты включают в себя только С-сегменты и фланкирующие их 5’и 3’-нетранслируемые участки. Количество стерильных транскриптов у сомика и форели в 5 раз превышает количество полных VJC-транскриптов.
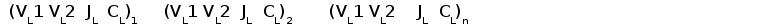




Рис. 5. Геномная организация локуса генов L-цепей ИГ у пещерного сомика.
Генные сегменты организованы в кластеры. Каждый кластер содержит два VL и по одному JL и CL сегменту и имеет размер около 5 тпн. Расстояние между JL и CL около 1000 пн, VL сегменты удалены от 5' конца JL сегмента на 3 тпн. VL сегменты всегда расположены в противоположной транскрипционной полярности по отношению JL и CL сегментам (Ghaffari and Lobb, 1993).
Амфибии.
У шпорцевой лягушки (Xenopus laevis) выявлено три семейства генов L-цепей ИГ, кодирующих три различных изотипа: L1 (ро), L2 (сигма) и L3 (рис. 6).
Все три локуса имеют сегментарный характер организации. L1 локус содержит множественные VL1 сегменты, разделенные в геноме 2.1 - 3.6 тпн, пять JL1 сегментов, три из которых идентичны, и один CL1 генный сегмент (Stewart et al., 1993). JL1 и VL1 сегменты фланкируются каноническими гепта- и нонамерными сигнальными последовательностями, разделенными 12 пн у VL1 и 23 пн у JL1 сегментов (Sakano et al., 1980).
Локус генов второго типа разделяется на два семейства, s1 и s2, которые в геноме располагаются отдельно. Геном лягушки содержит множественные Vs1 и несколько Vs2 геных сегментов, одну пару Js1-Cs1 и пару Js2-Cs2 (Schwager et al., 1991). Расположение генов этого типа друг относительно друга точно не определено.
Недавно был обнаружен третий тип генов L-цепей у лягушки. В геноме лягушки содержится шесть семейств V сегментов этого типа. Общее количество VL3 элементов составляет по меньшей мере 30 копий. Каждый VL3 элемент может рекомбинировать с одной из двух пар JCL3 генных сегментов (Haire et al., 1996).
Гены трех типов значительно различаются: гомология на аминокислотном уровне составляет приблизительно 30% (Zezza et al., 1991). В то же время, ряд признаков позволяет соотнести гены L1 и L3 типа с каппа и лямбда типами млекопитающих.
Семейство генов первого типа можно отнести к каппа типу по нескольким причинам (Zezza et al., 1992): а) высокая гомология первичной структуры (более 50%); б) существенное сходство геномной организации локуса (много VL1 и один CL1 генный сегмент); в) кофигурация спейсеров между гепта- и нонамерыми соответствует каппа типу; г) JL1 и CL1 генные сегменты разделены примерно 3.5 тпн, что приблизительно равно расстоянию между JLk и CLk элементами человека (Sakano et al., 1979; Hieter et al., 1982) и значительно больше чем расстояние между JLl и CLl млекопитающих (Blomberg et al., 1982; Udey, 1987).
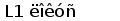

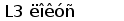
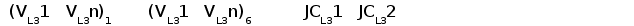


Рис. 6. Геномная организация локуса первого и третьего типов генов L-цепей ИГ у шпорцевой лягушки.
Локус семейства L1 содержит множественные VL1 сегменты, пять JL1 сегментов один CL1 генный сегмент. Расстояние между VL1 равно 2.1 - 3.6 тпн. Локус семейства L3 имеет около 30 VL3 сегментов, располагающихся группами, JL3 и CL3 сегменты располагаются парами: JCL31 и JCL32 (Stewart et al., 1993; Haire et al., 1996).
Третий тип генов L-цепей лягушки более близок к лямбда типу млекопитающих: а) более высокая гомология с генами лямбда типа млекопитающих (около 50% с l- и 30-40% с k-типом по аминокислотной последовательности); б) JL3 и CL3 генные сементы экспрессируются всегда парами JL31-CL31 и JL32-CL32, что напоминает ситуацию в лямбда локусе человека и мыши (Haire et al., 1996).
Гены второго типа L-цепей лягушки незначительное и приблизительно одинаковое сходство с генами каппа и лямбда типов млекопитающих (30-40% и 25-40% по нуклеотидной последовательности, соответствено) (Schwager et al., 1991).
Генетическое разнообразие у лягушки достаточно велико и сравнимо с разнообразием генов у млекопитающих. Однако репертуар антительных специфичностей в сыворотке крови значительно уступает репертуару высших позвоночных (Hsu et al., 1991). Этот парадокс можно объяснить наличием механизма соматической селекции клонов В-лимфоцитов, который элиминирует клетки, несущие антитела, специфичные к собственным антигенам организма, а также те клетки, которые продуцируют несколько типов антител (Wilson et al., 1992).
Основной вклад в формирование разнообразия L-цепей антител у лягушки дают первое и третье семейства генов, имеющие высокий комбинативный потенциал и большое разнообразие зародышевых V генных (Haire et al., 1996). Второе семейство имеет несколько слабо отличающихся друг от друга VL2 элементов и не играет существенной роли в формировании разнообразия (Stewart et al., 1993). Таким образом, разнообразие генов L-цепей у лягушки а создается посредством: а) комбинативного соединения V и J генных сегментов; б) смещения рекомбинационной рамки при соединении V и J сегментов. Нельзя исключить, что определенный вклад в разнообразие может вносить соматический мутагенез. Во всяком случае, наличие этого механизма показано для генов H-цепей лягушки (Zezza et al., 1992).
ЭВОЛЮЦИЯ ГЕНОВ L-ЦЕПЕЙ ИГ
Согласно наиболее популярной в настоящий момент гипотезе гены ИГ и Т-клеточных рецепторов произошли от одного предкового гена, кодировавшего один домен путем различных вариантов последовательных дупликаций (рис. 7).
В эволюции генов ИГ можно выделить три основных события.
1. Разделение предкового V-подобного гена на V и J сегменты и возникновение механизма генерации разнообразия путем соматической рекомбинации. Предпологается, что это событие связано с внедрением транспозона в предковый ген и осуществилось до разделения ИГ и Т-клеточных рецепторов у предков хрящевых рыб (Gilbert, 1990). Наличие в геноме акул и скатов слитых VJ генов в локусах L-цепей классов I и II, является, по-видимому, вторичным событием и, возможно, обусловлено фиксацией генов, кодирующих антитела строго определенной специфичности (Anderson et al., 1995).
2. Возникновение механизмов соматического мутагенеза генов ИГ. У млекопитающих соматический мутагенез обеспечивает как минимум половину разнообразия антител, являясь основой аффинного созревания антител при вторичном иммунном ответе (Пол, 1987). Достаточно выражен соматический мутагенез также у птиц (Thompson and Neiman, 1987; Reynaund et al., 1987).
Ряд данных позволяет предполагать, что соматический мутагенез может осуществляться у хрящевых рыб и у амфибий, но эти результаты нуждаются в более тщательном изучении. В частности, у хрящевых рыб получение достоверных свидетельств затруднено наличием множественых кластеров генов, что не позволяет надежно соотносить данные по геномной структуре и структуре кДНК (Litman et al., 1993).
3. Переход от кластерной к сегментарной организации генных сегментов. Сегментарная организация может иметь несколько преимуществ. Во-первых, увеличивается разнообразие продуцируемых антител за счет дополнительных возможностей комбинирования разных V и J сегментов. Во-вторых, уменьшается размер локуса за счет







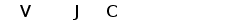
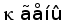
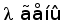



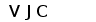

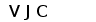

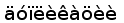



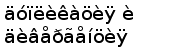
Рис. 7. Гипотетическая схема эволюции структурной организации локусов генов L-цепей ИГ позвоночных. (Anderson et al., 1995; Gilbert, 1990).
Обозначения:
- сигнальные
олигомеры,
фланкирующие
V и J сегменты.
резкого сокращения количества C генов. В-третьих, подобная организация позволяет упростить регуляцию экспрессии различных локусов.
Ключевым принципом функционирования иммунной системы является принцип клональной селекции иммунокомпетентных клеток, несущих рецептор определенной специфичности. Ряд косвенных данных позволяет считать, что у акул гуморальный иммунный ответ клонально ограничен (Shankey and Clem, 1980). Однако неизвестно, каким образом и насколько эффективно осуществляется это ограничение (Flajnik, 1996). У млекопитающих принцип “одна клетка - одно антитело” обеспечивается благодаря изотипическому и аллельному исключению. При сегментарной организации генов продуктивная перестройка в одном из локусов ведет к блокированию перестройки других локусов и, тем самым к их инактивации. Однако, в случае наличия множественных VJC кластеров, в части из которых VJ сегменты слиты в зародышевой ДНК, механизмы контроля выборочной экспрессии могут быть более сложными или не столь эффективными. Таким образом, переход от кластерной к сегментарной форме организации генов ИГ мог иметь принципиальное значение для формирования полноценной системы гуморального иммунитета.
Вопрос о том, на каком этапе филогенеза произошел этот переход остается открытым. Уже у амфибий гены ИГ L- и Н-цепей организованы сегментарно. При этом, в случае L-цепей наблюдается два основных типа организации: каппа-подобный (один С ген, семейство J сегментов и семейство V сегментов) и лямбда подобный (несколько пар JC и семейство V сегментов). По всей видимости, различные типы L-цепей амфибий произошли от различных типов L-цепей хрящевых рыб, однако относительно невысокая гомология по первичной структуре не позволяет утверждать этого с определенностью (Gilbert, 1990).
У сомика и радужной форели, представителей костистых рыб, организация генов L-цепей и Н-цепей отличается. Гены Н-цепей костистых рыб организованы подобно млекопитающим сегментарно (Bengten et al., 1991). Организация генов L-цепей имеет кластерный характер. В то же время, в отличие от хрящевых рыб, транскрипционная ориентация V генов у них изменена.
Следует отметить, что сомовые и лососевые являются довольно специализированной группой, возникшей в эволюции относительно недавно (Рис. 8). Особенности организации генов легких цепей у этих таксонов могут представлять собой новоприобретенные признаки (Romer, 1966).
В связи с этим особый интерес представляет изучение структры и организации генов L-цепей ИГ у костно-хрящевых рыб. Костно-хрящевые являются архаичной группой костистых рыб, возникшей значительно раньше настоящих костистых. Несмотря на то, что существуют различные мнения относительно последовательности появления костно-хрящевых и двоякодышащих (предков наземных позвоночных) (Ромер и Парсонс, 1992; Юдкин И. И., 1941), очевидно, что костно-хрящевые в большей степени чем коститстые сохранили черты древних первично-костных рыб и, соответственно, с большим основанием могут рассматриваться как промежуточное звено между хрящевыми рыбами и амфибиями.




















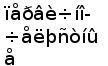


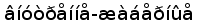


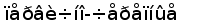

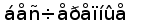
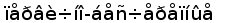
Рис. 8. Эволюционное древо низших позвоночных ( по Северцеву А. Н., 1939).
Обозначения:
- таксоны, имеющие
ныне живущих
представителей,
- таксоны,
существование
которых доказано
палеонтологическими
исследованиями.
МАТЕРИАЛЫ И МЕТОДЫ
ЭЛЕКТРОФОРЕЗ ДНК
В агарозном геле.
Электрофорез ДНК проводили в 1% агарозном геле в 1-кратном буфере ТАЕ, имеющем состав: 0.04 М трис-ацетат, 2 мМ Na2ЭДТА (Маниатис и др., 1984).
В полиакриламидном геле.
Электрофорез ДНК проводили в 6% полиакриламидном геле в 0,5-кратном буфере ТБЕ, имеющем состав: 0,089 М трис-борат, 0,089 М борная кислота, 2 мМ Na2ЭДТА. Для полимеризации геля к 50 мл раствора добавляли 300 мкл 10% раствора персульфата аммония и 30 мкл ТЕМЕД (Маниатис и др., 1984).
ВЫДЕЛЕНИЕ ДНК ИЗ ГЕЛЯ
Адсорбция на диэтиламиноэтилцеллюлозе.
Гель окрашивали в растворе бромистого этидия, участок геля с полосой ДНК вырезали, помещали в камеру и переносили ДНК на диэтиламиноэтилцеллюлозу в 0,5-кратном буфере ТБЕ (см. Электрофорез ДНК (2)) при напряжении 40 В/см. Элюцию ДНК с диэтиламиноэтилцеллюлозы проводили путем инкубирования в течение 30 мин при 70оС в буфере, имеющем состав: 2 мМ NaCl, 1 мМ Na2ЭДТА, 40 мМ трис рН 8,0 (Маниатис и др., 1984).
Адсорбция на кремниевой пудре.
Агарозный гель окрашивали в растворе бромистого зтидия, вырезали участок геля с ДНК и расплавляли инкубированием с двойным объемом 6 М раствора KI в течение 10 мин при 55оС. Затем добавляли суспензию кремниевой пудры (силики) в 3 М растворе KI в концентрации 100 мг/мл из расчета 300 мкг силики на 1 мкг ДНК и инкубировали в течение 2 мин при 55оС. После этого осаждали силику центрифугированием при 2000 g в течение 2 мин и дважды промывали суспендированием в растворе, содержащем 50 мМ NaCl, 10 мМ трис-HCl, рН 8,0, 2,5 мМ Na2ЭДТА и 50% этиловый спирт. Элюировали ДНК с силики суспендированием в воде (Boyle, Lew, 1995).
Замораживание - оттаивание.
Агарозный гель с ДНК 10 мин инкубировали в жидком азоте и осаждали центрифугированием при 12000 g в течение 15 мин, после чего добавляли к супернатанту 1/10 объема 3 М NaAc, 2 объема этанола и осаждали ДНК при 12000 g в течение 10 мин.
ПОЛУЧЕНИЕ РАДИОАКТИВНЫХ ЗОНДОВ
500 нг ДНК-матрицы и 100 нг праймера денатурировали в 10 мкл воды при 100оС в течение 10 мин и быстро охлаждали до 0оС. Затем добавляли 10 мкл 10-кратного буфера (40 мМ KP04 рН 7,5, 6,6 мМ MgCl2 и 1,0 мМ 2-меркаптоэтанол), 0,1-0,5 мКи 32Р-dATФ, 10 мкл 2 мМ раствора трех других немеченых дНTФ до концентрации 200 мкМ. Объем реакционной смеси доводили водой до 100 мкл, добавляли 3 мкл фрагмента Кленова ДНК-полимеразы I (10-12 е.а.) и инкубировали при комнатной температуре в течение 30 мин. Очистку зонда проводили методом гель - фильтрации на колонке с биогелем П-10 (Маниатис и др., 1984).
ВЫДЕЛЕНИЕ ПЛАЗМИДНОЙ ДНК
Клетки Escherichia coli, выращенные в среде Луриа-Бертани (1% NaCl, 1% бакто-триптон, 0,5% бакто-дрожжевой экстракт, рН 7,5) (среда LB) с ампициллином (50 мкг/мл), осаждали центрифугированием при 4000 g. Затем осадок суспендировали в буфере STET (8% сахароза, 0,5% тритон X-100, 10 мМ трис-HCl и 50 мМ Na2ЭДТА, рН 8,0) и добавляли лизоцим до концентрации 0,8 г/мл. Смесь инкубировали при комнатной температуре в течение 5 мин и при 100оС в течение 1 мин. После этого клеточный дебрис осаждали центрифугированием при 16000 g в течение 15 мин.
Для осаждения плазмидной ДНК к супернатанту добавляли 1/10 объема 3 М раствора ацетата натрия, равный объем изопропанола и центрифугировали при 16000 g в течение 15 мин при 20оС. Затем промывали осадок добавлением 80% этанола и повторным центрифугированием в течение 5 мин при 20оС (Маниатис и др., 1984).
ОЧИСТКА ПЛАЗМИДНОЙ ДНК МЕТОДОМ РАВНОВЕСНОГО ЦЕНТРИФУГИРОВАНИЯ В ГРАДИЕНТЕ ПЛОТНОСТИ CsCl
На каждые 10 мл раствора плазмидной ДНК, полученного после осаждения лизированных клеток (см. Выделение плазмидной ДНК), добавляли 1 г сухого CsCl, соль растворяли, после чего на каждые 10 мл раствора добавляли 0,8 мл раствора бромистого этидия с концентрацией 10 мг/мл. Смесь помещали в центрифужные пробирки Beckman, заполняли доверху раствором CsCl и бромистого этидия той же плотности и центрифугировали в роторе Ti 60 при 45000 об/мин и температуре 20оС в течение 36 часов. После этого отбирали кольцевую ковалентно замкнутую плазмидную ДНК и экстрагировали бромистый этидий из раствора до полного обесцвечивания равным объемом н-бутанола, предварительно насыщенного 1 М раствором NaCl.
ДНК осаждали центрифугированием при 15000 g, добавив равный объем 1 М раствора NH4Cl и два объема этанола. После промывания этанолом осадок ДНК растворяли в воде до концентрации 1 мкг/мкл (Маниатис и др., 1984).
ТРАНСФОРМАЦИЯ ПРОКАРИОТИЧЕСКИХ КЛЕТОК
Химическая трансформация.
Для подготовки компетентных клеток E.coli штамм XL-1 Blue MRF ' и выращивали в 100 мл 2-кратной среды Луриа (2% бакто-триптон, 1% бакто-дрожжевой экстракт, 0,1% NaCl, 0,4% глюкоза, рН 7,0) при 37оС до оптической плотности D550=0,35. Клеточную суспензию выдерживали при 0оС в течение 2 часов. Затем клетки осаждали центрифугированием при 4000 g, температуре 4оС и суспендировали в 50 мл ледяного буфера А (100 мМ CaCl2, 70 мМ MnCl2, 40 мМ NaAc, рН 5,5). После этого суспензию клеток выдерживали при 0оС в течение 30 мин, после чего клетки осаждали и суспендировали в 5 мл буфера А с 15% глицерина.
Для трансформации к 200 мкл суспензии компетентных клеток добавляли 30 нг плазмидной ДНК, растворенной в 100 мкл воды и выдерживали при 0оС в течение 30 мин. Затем инкубировали при 37оС в течение 5 мин, после чего добавляли 3 мл среды LB и инкубировали при 37оС еще 90 мин. Клетки собирали центрифугированием и высевали на селективную среду (Мазин и др., 1988).
Электротрансформация.
Для подготовки компетентных клеток E.coli штамм XL-1 Blue MRF ' выращивали в 100 мл среды LB (см. Химическая трансформация) при комнатной температуре до оптической плотности D550=0,45. Затем клетки осаждали центрифугированием при 4000 g, 4оС и промывали суспендированием последовательно в 100 мл, 50 мл и 10мл 10% раствора глицерина при 0-4оС. После этого клетки суспендировали в 240 мкл ледяного раствора GYT (10% глицерин, 0,125% бакто-дрожжевой экстракт, 0,25% бакто-триптон).
Для трансформации к 40 мкл суспензии компетентных клеток добавляли 1 мкл ДНК вектора (50 нг) при 0-4оС и пропускали электрический разряд (U=18000 В). Затем добавляли 1 мл SOC (2% бакто-триптон, 0,5% бакто-дрожжевой экстракт, 10 мМ NaCl, 2,5 мМ KCl, 10 мМ MgCl2, 10 мМ MgSO4, 20 мМ глюкоза), перемешивали, переносили в 3 мл среды SOC, прогретой до 42оС, и инкубировали при 37оС в течение 1 часа (Wai Lin Tung and King-C. Chow, 1995).
ВЫДЕЛЕНИЕ ЛЕЙКОЦИТОВ ИЗ КРОВИ РЫБ
Кровь собирали на холоду через каудальную артерию с добавлением на каждые 5 мл крови 1 мл 2,7% раствора Na2ЭДТА, рН 7,4. К выделенной крови добавляли равный объем трис-солевого буфера (TBS) (0,14 М NaCl, 5 мМ KCl, 25 мМ трис-HCl, рН 7,4). Смесь наносили при комнатной температуре на равный объем среды для выделения лейкоцитов, содержащей 24 части 9% фикола и 10 частей 34% верографина. После центрифугирования при 400 g в течение 30 мин отбирали слой лейкоцитов и разводили в большом объеме буфера TBS. После повторного центрифугирования при 300 g в течение 40 мин для лизиса остаточных эритроцитов клетки суспендировали в растворе, содержащем 9 объемов 0,83% раствора NH4Cl и 1 объем 2,06% раствора трис-HCl, рН 7,62, до концентрации 108 кл/мл и добавляли 5 объемов среды. После осаждения клеток при 300 g в течение 10 мин процедуру очистки повторили (Фримель, 1987).
ВЫДЕЛЕНИЕ СУММАРНОЙ РНК ИЗ ЛЕЙКОЦИТОВ
Осадок лейкоцитов после выделения и очистки суспендировали в 10 объемах (1 мл) раствора 1, содержащего 4 М гуанидин тиоцианат, 25 мМ цитрат натрия, рН 7,0, 0,5% саркозил и 0,1 М 2-меркаптоэтанол, при комнатной температуре. Затем добавляли 0,1 мл 2 М раствора NaAc, рН 4,0, 1 мл фенола, 0,2 мл хлороформа и перемешивали. Затем центрифугировали при 10000 g в течение 20 мин при 4оС, отбирали водную фазу, добавляли к ней равный объем изопропанола и осаждали РНК центрифугированием при 15000 g в течение 15 мин при 4оС (Chomczynski and Sacchi, 1987).
ВЫДЕЛЕНИЕ ВЫСОКОМОЛЕКУЛЯРНОЙ ДНК ИЗ ЭУКАРИОТИЧЕСКИХ КЛЕТОК
1 г печени стерляди, замороженной в жидком азоте, растирали в тонкий порошок и гомогенизировали в 15 мл раствора, содержащего 0,5 М Na2ЭДТА, рН 8,0, 0,5% саркозила, 100 мкг/мл протеиназы К и нагретого до 65оС. Затем инкубировали при 50оС течение 3 часов при легком покачивании. Затем, избегая резких встряхиваний, экстрагировали ДНК равным объемом фенола три раза, каждый раз разделяя фракции центрифугированием при 4000 g и комнатной температуре. Фенол предварительно очищали перегонкой и насыщали буфером, содержащим 1 М трис-HCl, один раз и два раза буфером, содержащим 0,1 М трис-HCl и 0,1% 2-меркаптоэтанол. Диализ ДНК проводили при 10оС в течение 12 часов против 4 л буфера, имеющего состав:
50 мМ трис-HCl, рН 8,0,
10 мМ Na2ЭДТА,
10 мМ NaCl.
Затем обрабатывали пробу РНКазой, свободной от ДНКазы,при 37оС в течение 1 часа в концентрации 100 мкг/мл. После этого экстрагировали ДНК равным объемом смеси фенол-хлороформ (1:1) два раза и проводили диализ при 10оС в течение 36 часов против 10 л буфера TE (10 мМ трис-HCl, 1 мМ Na2ЭДТА, рН 7,5), меняя буфер через каждые 12 часов (Маниатис и др., 1984).
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК
Выделение поли(A)+РНК.
Для ингибирования РНКазы все растворы, не содержащие трис, обрабатывали 0,1% диэтилпирокарбонатом 12 часов при 37оС и автоклавировали. Посуду прокаливали при 250оС в течение 4 часов. Поли(A)+РНК выделяли из суммарной РНК лейкоцитов стерляди методом хроматографии на олиго(dT)-целлюлозе. Для этого РНК растворяли в воде, прогревали при 65оС в течение 5 мин, добавляли равный объем 2-кратного буфера для нанесения РНК (40 мМ трис-HCl, рН 7,6, 1 М NaCl, 2 мМ Na2ЭДТА, 0,2% SDS) и трижды пропускали через колонку с олиго(dT)-целлюлозой. Затем промывали колонку 5-10 объемами буфера для нанесения и 4 объемами этого же буфера, содержащего 0,1 М NaCl. После этого элюировали поли(A)+РНК в 2-3 объемах воды, обработанной диэтилпирокарбонатом, добавляли 2,2 объема этанола и осаждали центрифугированием в течение 10 мин при 12000 g при 4оС (Маниатис и др., 1984).
Синтез кДНК.
Синтез кДНК проводили в соответствии с протоколом фирмы Stratagene. По матрице поли(A)+РНК синтезировали первую цепь кДНК (рис. 9).
Реакционная имела состав:
5 мкл 10-кратного буфера для синтеза первой цепи,
3 мкл смеси метил-dНTФ,
2 мкл раствора праймер-линкера (1.4 мкг/мкл),
1 мкл ингибитора РНКазной активности,
1.5 мкл фермента обратная транскриптаза (50 е.а./мкл),
вода до суммарного объема 50 мкл.
Смесь инкубировали при 37оС в течение 1 часа. Затем охлаждали до 0оС и добляли компоненты для синтеза второй цепи кДНК:
20 мкл буфера для синтеза второй цепи,
6 мкл смеси dНTФ,
88,9 мкл стерильной воды,
2 мкл 32Р-dНTФ (800 Ки/мМ),
2 мкл раствора РНКазыH (1.5 е.а./мкл),
11.1 мкл ДНК полимеразы I.
Смесь инкубировали при 16оС в течение 2,5 часов, затем охлаждали до 0оС и добавляли компоненты для застройки “липких” концов:
23 мкл смеси дНTФ,
2 мкл Pfu ДНК полимеразы (2,5 е.а./мкл).
Смесь инкубировали при 72оС в течение 30 мин.
Затем проводили экстракцию равным объемом смеси фенол-хлороформ (1:1) и равным объемом хлороформа, после чего отбирали водную фазу, добавляли 20 мкл 3М NaAc и 400 мкл 96% этанола и осаждали кДНК центрифугированием при 12000 g в течение 12 мин.
Сшивка молекул кДНК с вектором.
кДНК растворяли с EcoR I адапторами (рис. 9) и инкубировали при 4оС в течение 30 мин. Затем добавляли компоненты для сшивки молекул адапторов и молеул кДНК:
праймер-линкер
5' CTCGAGTTTTTTTTTTTT 3'
 3' AAAAAAAAAAAA 5'
3' AAAAAAAAAAAA 5'
 поли(A)+РНК
поли(A)+РНК
Xho I сайт 1-я цепь EcoR I адаптор


 5' р-TCGAGTTTTTTTTTTTT,,,,,,,,CTCGTGCCG-р
3'
5' р-TCGAGTTTTTTTTTTTT,,,,,,,,CTCGTGCCG-р
3'
3' р-CAAAAAAAAAAAA
GAGCACGGCTTAA-р 5'
 2-я
цепь
2-я
цепь



Xho I сайт кДНК EcoR I сайт

 5' CTCGAGTTTTTTT,,,,,,,,,CTCGTGCCGAATTC 3’
5' CTCGAGTTTTTTT,,,,,,,,,CTCGTGCCGAATTC 3’
3' GAGCTCAAAAAAA GAGCACGGCTTAAG 5’
ДНК вектора ДНК вектора
Рис. 9. Схема синтеза кДНК по матрице поли(A)+РНК и сшивки с плазмидным вектором
pBluescript KS(+) по сайтам EcoR I и Xho I.

 Обозначения:,,,,,
- метилированная
цепь кДНК,
-неметилированная
цепь кДНК.
Обозначения:,,,,,
- метилированная
цепь кДНК,
-неметилированная
цепь кДНК.
1 мкл 10-кратного буфера лигирования,
1 мкл 10 мМ рATФ,
1 мкл фермента ДНК лигазы фага Т4 (4 е.а./мкл).
Смесь инкубировали при 4оС в течение 2-х суток, после чего инактивировали лигазу прогреванием при 70оС в течение 30 мин, охлаждали до комнатной температуры и добавляли компоненты для фосфорилирования концевых нуклеотидов:
1 мкл 10-кратного буфера лигирования,
2 мкл 10 мМ рATФ,
6 мкл стерильной воды,
1 мкл фермента полинуклеотидной киназы фага Т4 (10 е.а./мкл).
Реакционную смесь инкубировали при 37оС в течение 30 мин. Затем охлаждали до комнатной температуры и добавляли компоненты для образования липких концов по сайту рестрикции эндонуклеазы Xho I:
28 мкл буфера 4,
3 мкл Xho I (40 е.а./мкл).
Смесь инкубировали при 37оС в течение 1,5 часа. Затем добавляли 5 мкл 10-кратного буфера STE (1 М NaCl, 200 мМ трис-HCl, рН 7,5, 100 мМ Na2ЭДТА) и пропускали через колонку с гелем сефакрил С-500. Фракции, содержащие молекулы с размером более 500 пн объединяли и экстрагировали ДНК равным объемом смеси фенол-хлороформ (1:1) и равным объемом хлороформа. Затем добавляли двойной объем этанола, осаждали кДНК центрифугированием, промывали 80% этанолом и растворяли в 10 мкл воды. Для лигирования брали 5 мкл раствора кДНК (600 нг) и 2 мкл раствора вектора pBluescript KS(+) (1 мкг), имеющего "липкие" дефосфорилированные концы по сайтам рестрикции эндонуклеазами EcoR I и Xho I (рис. 9).
Затем добавляли 1 мкл 10-кратного буфера 3, 1 мкл 10 мМ раствора рATФ и 1 мкл фермента лигазы фага Т4 (4 е.а./мкл). Смесь инкубировали при 4оС в течение 2-х суток, после чего экстрагировали равными обьемами фенол-хлороформа и хлороформа.
Амплификация библиотеки.
Электротрансформацию проводили как описано в п. Трансформация прокариотических клеток, после чего высевали 1/1000 объема (2,5 мкл) среды SOC на селективную среду с ампициллином (50 мкг/мл) для определения количества трансформированных клеток. Остальную часть помещали в 100 мл среды LB с ампициллином (50 мкг/мл) и инкубировали при 37оС при перемешивании до тех пор, пока оптическая плотность не достигнет величины D550=2-2,5. Затем отбирали 1,6х1011 клеток, осаждали их центрифугированием, суспендировали в 4 мл среды LB и перемешивали с 4 мл глицерина. Полученную библиотеку хранили при -70оС.
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Реакционная смесь полимеразной цепной реакции (ПЦР) содержала следующие компоненты:
100 нг ДНК матрицы,
100 нг невырожденного праймера,
500 нг вырожденного праймера,
5 мкл 2 мМ раствора дНTФ,
5 мкл 1-кратного буфера (10 мМ трис-HCl, рН 8,8, 50 мМ KCl, 1,5 мМ MgCl2, 0,001% желатин),
1 мкл Taq полимеразы,
вода до конечного объема 50 мкл (Dieffenbach and Dveksler, 1995).
Температурный режим ПЦР.
Реакционную смесь инкубировали при 95оС в течение 5 мин, после чего добавляли Taq полимеразу и использовали температурный режим, при котором в течение первых 18 циклов температуру отжига праймеров понижали на 3оС черех каждые три цикла:
1й-18й циклы 95оС - 1 мин (денатурация ДНК),
50-35оС - 1 мин (отжиг праймеров),
72оС - 2 мин (полимеризация).
19-й-30-й циклы: 95оС - 1 мин,
50оС - 1 мин,
72оС - 2 мин.
31-й цикл: 95оС - 1 мин,
50оС - 1 мин,
72оС - 10 мин.
СУБКЛОНИРОВАНИЕ ДНК
Фрагменты ДНК, полученные в результате ПЦР, разделяли в 1% агарозном геле и выделяли как описано в п. Выделение ДНК из геля. Затем осаждали ДНК этанолом, растворяли в 5-10 мкл воды и добавляли компоненты для удаления выступающих нуклеотидов с 3' конца ДНК:
1 мкл 10-кратного буфера (200 мМ трис-HCl, рН 8,2, 100 мМ KСl, 60 мМ (NH4)2SO4, 20 мМ MgCl2, 1% тритон Х-100 и 100 мкг/мл бычий сывороточный альбумин),
1 мкл 10 мМ dНTФ,
1 мкл Pfu ДНК полимеразы (2,5 е.а./мкл),
воду до конечного объема 10 мкл.
Смесь инкубировали 30 мин при 72оС после чего охлаждали до 0оС и добавляли компоненты для сшивки с вектором pBluescript KS(+) по сайту рестрикции Sma I:
1 мкл расщепленного вектора (0,5 мкг),
1 мкл 10-кратного буфера (50 мМ трис-HCl, рН 7,5, 7 мМ MgCl2, 1 мМ ДТТ),
1 мкл 10 мМ дATФ,
1 мкл ДНК лигазы фага Т4 и воду до конечного объема 10 мкл.
Смесь инкубировали одни сутки при комнатной температуре и затем использовали для химической трансформации клеток E.coli. Трансформированные клетки высевали на селективную среду: 1,5% LB-агар с ампициллином (50 мкг/мл), тетрациклином (15 мкг/мл), ИПТГ (2 мМ), X-Gal (500 мкг/мл) и инкубировали 14 часов при 37оС. После этого отбирали колонии белого цвета (Маниатис и др., 1984; Dieffenbach and Dveksler, 1995).
СКРИНИНГ БИБЛИОТЕКИ кДНК
Библиотеку кДНК высевали на чашки Петри с 1,5% LB-агаром с ампициллином (50 мкг/мл) из расчета 4000 колоний на чашку и инкубировали при 37оС до тех пор пока колонии не достигнут 0,5-1 мм в диаметре. Колонии переносили на нейлоновую мембрану и инкубировали при комнатной температуре в денатурирующем растворе (0,5 М NaOH, 1,5 М NaCl) в течение 7 мин и два раза по 2 мин в нейтрализующем растворе (1,5 М NaCl, 0,5 М трис-HCl, рН 7,5). Затем споласкивали мембрану в 2-кратном буфере SSC (0,3 М NaCl, 0,03 М цитрат натрия), высушивали и фиксировали ДНК в течение 10 мин ультрафиолетовым излучением с длиной волны 310 нм. После этого мембрану помещали в 50 мл предгибридизационного раствора (раствор 1), содержащего:
6-кратный SSC (0.9 М NaCl, 0,09 М цитрат натрия),
5-кратный раствор Денхардта (0,1% бычий сывороточный альбумин, 0,1% фикол, 0,1% поливинилпиролидон),
0,5% SDS,
10% декстран сульфат,
500 мкг денатурирированной ДНК цыпленка,
и инкубировали 1 час при 55оС. После этого добавляли меченую 32P-дATФ ДНК зонда и инкубировали 12-18 часов при 55оС при постоянном покачивании.
После гибридизации проводили промывание мембраны два раза по 10 мин в 200 мл 2-кратного SSC + 0,1% SDS при комнатной температуре, 15 мин в 1-кратном SSC + 0,1% SDS% при 55оС и два раза по 10 мин в 0,1-кратном SSC + 0,1% SDS при 55оС. Экспонирование проводилось в течение 12 часов при -70оС с рентгеновской пленкой (Маниатис и др., 1984).
ОПРЕДЕЛЕНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК
Нуклеотидную последовательность ДНК определяли по методу Сэнгера (Sanger et al., 1977). 5 мкг плазмидной ДНК, очищеной гель-фильтрацией на колонке с биогелем П-10, денатурировали в 50 мкл 0.2 М NaOH при 37оС в течение 30 мин, затем добавляли 5 мкл 3 М NaAc, рН 5,1, двойной объем этанола и инкубировали 30 мин при температуре -70оС. После этого осаждали ДНК центрифугированием при 12000 g в течение 12 мин и промывали осадок 100 мкл 80% этанола. Осадок ДНК высушивали при комнатной температуре и растворяли в 7,5 мкл воды, добавляли 2 мкл 5-кратного буфера (200 мМ трис-HCl, рН 7,5, 100 мМ MgCl2, 250 мМ NaCl), 0,5 мкл праймера (50 нг) и инкубировали 30 мин при 37оС. После этого пробирку помещали в лед и добавляли 1 мкл 0,1 М ДТТ, 2 мкл 1,5 мкМ дНTФ без дATФ, 0,5 мкл раствора 35S-дATФ и 2 мкл ДНК полимеразы фага Т7 (2,5 е.а./мкл). Затем инкубировали 3 мин при 20оС и сразу отбирали по 3,5 мкл в четыре пробирки, каждая из которых содержала по 2,5 мкл 80 мкМ смеси четырех дНTФ, 50 мМ NaCl и 8 мкМ одного из ддНTФ. Инкубировали 5 мин при 37оС и добавляли по 4 мкл стоп-раствора (95% формамид, 20 мМ Na2ЭДТА, 0,05% бромфеноловый синий, 0,05% ксилен цианол). Полученные образцы инкубировали 3 мин при 100оС, быстро охлаждали до 0оС и отбирали по 2 мкл для электрофореза в денатурирующем полиакриламидном геле, имеющем состав:
6% акриламид,
8 М мочевина,
1-кратный глицерин-толерантный буфер (1,1% трис, 0,36% таурин, 0,02% Na2ЭДТА).
Электрофорез проводили при напряжении 40 В/см. Затем гель выдерживали 30 мин в 10% уксксной кислоте, высушивали и экспонировали 12 часов при комнатной температуре с рентгеновской пленкой (Маниатис и др., 1984).
САУЗЕРН БЛОТ ГИБРИДИЗАЦИЯ
10 мкг геномной ДНК, выделенной из печени стерляди, инкубировали в течение 2 часов с эндонуклеазами рестрикции Pst I и Pvu II (по 50 е.а.) при 37оС в буфере, содержащем 100 мМ NaCl, 10 мМ трис-HCl, рН 7,5, 10 мМ MgCl2, 100 мкг/мл бычий сывороточный альбумин, 6 мМ 2-меркаптоэтанол. Затем экстрагировали ДНК последовательно равными объемами фенол-хлороформа (1:1) и хлороформа, осаждали ДНК двумя объемами этанола, растворяли в воде и наносили на 1% агарозный гель. Электрофорез проводили при напряжении 1 В/см и температуре 12оС в течение 12 часов. Затем гель инкубировали при комнатной температуре 30 мин в 0,25 М HCl, 30 мин в денатурирующем растворе (0,5 М NaOH, 1,5 M NaCl) и 10-15 мин в растворе 1 (0,25 М NaOH, 1,5 M NaCl). После этого ДНК переносили на нейлоновую мембрану методом вакуумного переноса в растворе 1. Мембрану споласкивали в 2-кратном SSC, высушивали и фиксировали ДНК в течение 10 мин ультрафиолетовым излучением с длиной волны 310 нм.
Предгибридизацию проводили при 55оС в течение 1 часа в растворе 1 (см. Скрининг библиотеки кДНК). Гибридизация длилась 12-16 часов при 55оС с концентрацией зонда не более 10 нг/мл.
Отмывку проводили два раза по 10 мин в 2-кратном SSC + 0,1% SDS при комнатной температуре и один раз 15 мин в 1-кратном SSC + 0,1% SDS при 55оС. Экспонирование длилось 12 часов при -70оС (Маниатис и др., 1984).
РЕЗУЛЬТАТЫ
КОНСТРУИРОВАНИЕ БИБЛИОТЕКИ кДНК И ЕЕ СКРИНИНГ
В качестве источника мРНК брали лейкоциты периферической крови стерляди. На основе этой мРНК синезировали кДНК. Полученные молекулы кДНК клонировали в плазмидном векторе pBluescript KS(+), после чего трансформировали этим вектором клетки E.coli штамма XL-1 Blue MRF’. В результате этого была получена библиотека кДНК представительностью 6 миллионов клонов. Библиотеку амплифицировали до объема 5 х 1011 клеток.
Cуммарную плазмидную ДНК библиотеки выделяли и использовали в ПЦР с парой праймеров, один из которых гомологичнен участку вектора вблизи сайта клонирования молекулы кДНК и располагается в кодирующей ориентации. Второй праймер вырожденный, соответствующий наиболее консервативному участку последовательности J-сегментов L-цепей ИГ позвоночных в некодирующей ориентации (рис. 10).
Один из фрагментов ДНК, полученных в результате ПЦР, соответствовал ожидаемому размеру (370 пн). Этот фрагмент выделяли и субклонировали в векторе pBluescript KS(+). Определение нуклеотидной последовательности этого фрагмента показало, что он гомологичен генам V области L-цепей ИГ (Рис. 11).
Этот фрагмент был использован в качестве зонда для скрининга библиотеки кДНК лейкоцитов стерляди. Скрининг 4000 колоний библиотеки кДНК в мягких условиях гибридизации позволил выявить пять клонов с размером вставки кДНК от 800 до 1050 пн. Для четкого разделения клонов проводили вторичный скрининг библиотеки кДНК, после чего выделенные клоны амплифицировали.
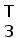


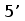
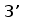







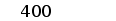
прамер Т3
5'-GCAATTAACCCTCACTAAAGG-3';
J-праймер
5'-A(G/C)(T/C)(T/C)TGGT(T/G/C)CCTTTTCC(A/G)AA-3’
Рис. 10. Схема ПЦР. Т3 праймер располагается вблизи сайта клонирования молекулы кДНК и комплиментарен некодирующей цепи, J-праймер гомологичен участку кодирующей цепи J-сегмента генов L-цепей ИГ позвоночных. Ожидаемая длина продуктов около 400 пн.
ОПРЕДЕЛЕНИЕ ПЕРВИЧНОЙ СТРУКТУРЫ
Определена нуклеотидная последовательность вставки кДНК клона В1. кДНК клона В1 размером 1050 пн представляла собой полноразмерную копию зрелого гена L-цепи ИГ и содержала 5’-нетранслируемую область, последовательность кодирующую лидерный пептид, V область, С область, а также 3’-нетранслируемую область (Рис. 11). На основе первичной структуры клона В1 сконструировали и синтезировали три праймера, соответствующих консервативным районам V и C области и использовали их для определения нуклеотидной последовательности клонов Е4 и С2. Последовательность клона Е4 была определена только в области, примыкающей к Т3 промотору вектора. За исключением 2 нуклеотидов она полностью идентична последовательности В1. Последовательность клона С2 имела химерный характер. В области Т3 промотора она содержала фрагмент соответствующий 3’ части С сегмента клона B1 с заменами нескольких нуклеотидов. В то же время использование для определения последовательности праймеров V и C области показало, что эта вставка содержит дополнительно перестроенный ген легкой цепи, содержащий большую часть V сегмента и второй С сегмент, отличающийся от первого в одной позиции. Поскольку пока не удалось определить последовательность в области стыка этих двух фрагментов вставки, нельзя сказать, находятся ли они в одной или разных ориентациях. По-видимому, сходный химерный характер имеет последовательность клона D2 (данные не показаны).
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ
Нуклеотидную и предсказанные аминокислотные последовательности клона В1 сравнивали с последовательностями V и C генов L-цепей ИГ других видов позвоночных (Рис 12). На аминокислотном уровне V область имеет наибольшее сходство с V-областями L-цепей класса F и G пещерного сомика и с V-областью L1 класса ИГ форели (63-71%). И те и другие относятся к каппа-типу L-цепей. При поиске гомологов в полном банке последовательностей ИГ млекопитающих первые 100 позиций с наибольшим сходством представляли собой также V каппа области.
С-область клона В1 имеет на аминокислотном уровне приблизительно одинаковое и не очень значительное сходство с широким спектром С-областей L-цепей ИГ хрящевых и костистых рыб (41-44%). На нуклеотидном уровне наиболее близкими гомологами В1С гена являются гены III класса хрящевых рыб (65%).
АНАЛИЗ ГЕНОМНОЙ ОРГАНИЗАЦИИ
Геномную организацию локуса генов L-цепей ИГ стерляди оценивали методом Саузерн блот гибридизации. Геномную ДНК выделяли из печени стерляди. ДНК расщепляли эндонуклеазами рестрикции Pst I и Pvu II. После электрофоретического разделения и переноса фрагментов ДНК на нитроцеллюлозную мембрану проводили гибридизацию в мягких условиях. В качестве зонда на V-гены использовали фрагмент, полученный в результате ПЦР (см. Конструирование библиотеки кДНК и ее скрининг). В качестве С-зонда использовали участок кДНК клона В1, кодирующий CL сегмент, который вырезали по сайтам рестрикции эндонуклеазами EcoR V-Xho I.
V-зонд выявлял более 20 гибридизующихся фрагментов ДНК, в то время как С-зонд гибридизовался только с 3-4 фрагментами (рис. 13).
5' нетранслируемая область |> лидерный пептид
1 15 30 45 60 75 90
B1 . .TTCATGGCAACA GAATCCACAACAACA ATGACTTTTATCAGC ATCTTCATCTGGGCA CTTGTCATCTGCACT CAGGAATCCAGTGGA
E4 CCC----A------- --------------- --------------- --------------- --------------- ---------------
C2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
|> V сегмент FR 1 |> CDR 1
105 120 135 150 165 180
B1 CAGTATACTGTGACT CAGACTCCAGCAGAG AAATCTGTTCTCCCA GGAGACACAGTCGCT CTCAACTGTAAAGTC AACAGCGCAGTCCTT
E4 --------------- --------G----T- --------------- ----------. . . . . . . . . . . . . . . . . . .
C2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
|> FR 2 |> CDR 2 |> FR 3
195 210 225 240 255 270
B1V GGTAATACGTACCTG CACTGGTACCAACAG AAACCTGGAGAAGCT CCCAAACTCCTGATC TATAGGGCAAGTACC CTTGAGTCTGGGATC
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ---- ---C-----------
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PCR . . . . . . . . . . . . . . . . . .------------ -----------A--- ---TAT----T--AG ---C-------N---
285 300 315 330 345 360
B1 CCAACTCGTTTCAGT GGCAGTGGATCTGGG ACTGACTTCACTCTG ACCATCAGTGGAGTC CAGGCTGAAGATGAA GGAGATTACTACTGT
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 --------C----C- --------------- --------------- --------------- --------------- --------T------
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PCR --------------- ------------TT- --------------- --------------- -------------C- ---------------
|> CDR 3 |> J сегмент |> С сегмент
375 390 405 420 435 450
B1 GTCAGTGTACACTAC TCCAGCAGTAGATAT GTGTTGACTTTCGGG CCCGGGACCAAACTA ATTGTGAAATCTGGA AGCCCCACTGCTCCT
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 CAG---TC------- C----------C-G- -NCC-C--------A --A--------G--G G-------------- -A-------------
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
PCR CAG---CAC---ACT C---ATG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
465 480 495 510 525 540
B1 TCCTCCGTCTCCCTG CTTCCTCCCTCCAAG CTGGAGCTCGACTCA AAGGGTAAAGCCACC CTGGTCTGCCTGGTG AATAATTTCTACCCT
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 --------------- --------------- ------------A-- -----C--------- TG------------- --------------C
C2(T3) . . . . . . . . . . . . . . . . . . . . . . . . . . -C--------- T-------------- --------------C
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
555 570 585 600 615 630
B1 GATGTCGTGGATATC AAGTGGACGGTGGAC GGGGTGGCCCAGTCG TCTGGTGTGCTGACC AGCACAATGAAGCAG AAGGATGGGAAATAC
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 --------------- --------------T ----C---------- --------------- --------------- -----C---------
C2(T3) --------------- --------------T ----C---------- --------------- --------------- -----C---------
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
645 660 675 690 705 720
B1 AGTGCAAGCAGCAGC CTGACCCTCACCAAG GCCGTGTGGAACAGC AAGGAGACATACACC TGCACTGTGAAGCAC GAGGCTGTGAGCACT
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 --------------- --------------- -A------------- --------------- --------------- ---------------
C2(T3) --------------- --------------- -A------------- --------------- --------------- ---------------
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
|> 3' нетранслируемая область
735 750 765 780 792
B1 CCCAGGAGCGAGTCC ATCAAGAGGAGCGAG TGCACACTATTAGAT GCCtaaAGG . . . . . . . . .
E4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C2 --G----------T- --------------- --------------- ---taaG-CAAGAGA ATCCGTGTGATT
C2(T3) --G----------T- --------------- --------------- . . . . . . . . . . . . . .
PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Рис. 11. Нуклеотидная последовательность полноразмерной кДНК клона В1 и частичная последовательность кДНК клонов Е4 и С2 и продукта ПЦР. Последовательность кДНК клона С2 определялась с использование праймеров V1, V2, C1 (обозначение С2) и Т3 (обозначение С2(Т3)). Прочерк означает идентичность в этой позиции с кДНК клона В1, FR - каркасные участки, CDR - гипервариабельные участки V области.
A
AruIgLBV QYTVTQTPAEKSVLPGDTVALNCKVNSAVLGNTYLHWYQQKPGEAPKLLIY*RASTLESGIPTRFSGSG**SGTDFTLTISGVQAEDEGDYYCVSV
IpuIgLVG -V-------V--A---E--TI--RT-P-Y*-H*-----------------K*F-NQ-H----A------**--S----------T--A-----Q-Y 71%
OmyIgLV1 -I------EM-AFQT--A-T-R-RF-KPSPPC**VA-------G--Q----*Y-T--Q--T-S------**--S-------------A-----Q-Y 63%
XIaIgLVR -VVL--S-DYV--S--E--TIT--AS-SS---**-------S-QT------*GT-NRYT-T-E------**----------RME---AA----QQY 58%
Rabbit kV IV ivm----ss---pv----ti--qasqs-ys-nr-a-f-----qp------*k----a--v-s--k---**---q------d--ca-aat---rva 57%
GciIgLVIII -M--S-PVL--GL-Q--TIT-TASQS-YS-**-A----RE-QK-S----*A-TNRYTEVSE------**---S-----RN--P--VA----QGT 48%
HulV2 -SAL--*-ASV-GS--QSITIS-TGT-SSYNL**VS----H--K----M--*EG-KRP--VSN-----K**--NTAS--T--L-----A----C-Y 47%
HfrIgLVII GTVL--*--SM-TSQ-K--KIT-TISGGGTYY**SS--W----S--VFVWRDYD--RG----D--T--RNT-SNVMH---TD--SR-TA-----AW 41%
XlaIgLIII -VSI--*-VSE--KL-E--RIS-TLSG-SGYH**VN-----A-NR-RY-LRFYSDSNK**HQD-----KDSPNNIGY---K-ALL--DA----ATW 35%
HfrIgLV1 VPVLN---ISDP-SA-E-SE-K-AMQNGGSYY**MS--R-R-----VFVL**YQ--SG-IYRD--KP-RDT-SNSHI---GSLEPG-SAV---AAN 31%
Б
AruIgLCB GSPTAPSSVSLLPPSKLELDSKGKATLVCLVNNFYPDVVDIKWTVDGVAQSSG*VLTSTMKQK**DGKYSASSSLTLTKAVWNSKETYTCTVKHEAVSTPRSESIKRSECTLLDA
CplIgLCII -RS*PT--V----SDQITA-NM-------SG-V-GAAE-E-----SVRGN-*-E--RIQ-EA*-NTF-V--Y---SASD---H-L-S-V-K-ETQAN-LQT--S--S-M 44%
HfrIgLCIII EKSQPTLT-M---PE-VKA--T------ADH----E-GVE-KK--A-I-A-*-Q--NYLRAS*-ST--C--L---SGSD-E-NARFS-ALT-ETL-S-L-K-VS----V 42%
IpuIgLCF G--VKP-----L--S-Q-S*E-S-S-L--LPAYS-QGALVS-----SEVKD-*----AEER-**TDG-TR--T---S--L-EKG-EFV-K-S-DN-DH-VT**FRK-Q-EV 41%
Hu kappa TV-AP-VFIF---*D-QLKS-T-SV---L-----REAKVQ-K--NAL--GN*SQE-VTE-DSK-ST--L--T---S--DYEKHKV-A-E-T-QGL-S-VTK-FN-G-- 39%
XlaIgLC3 GDVK--*---YF---*V-EIATK---V--SLSD-T-RGATVK-L---KD-TDS*-QS-GLSKQS*-NL-ME--Y-S--ADQ-LRH---S-K-S-Q**GKEIIQTL-----V 38%
Hu lambda1 Q-K-NPT-T-F---*S-ELQAN-------ISD---GA-TVA-KA--SPVKA-*-E-TKPSKQS*NN--A---Y-S--PEQ-K-HRS-S-Q-T-E**GSTVEKTVAPT--S 37%
HfrIgLCI SEDRKP-VL-----*S-EIDS-W---S---SR-K-GF-RVL-R--DKETD--*-T-G-VSTDS*-QS--L--YLRVPATA--KGSS---S-D-GSL-S-LLKT-SSTA-SD 37%
XlaIgLCS ND-KPA-FIFK--*D-QVKE-NP-A---I---F-RDLTVT-K--SQDV--SD-K--DFM-ES*-ST--Q--M-T---DK-DKADKFE-L-K-K**TAQLTQSFSK-Q-S 34%
IpuIgLCG LTQP--TV----SV--Q*QE-V-----AYKGF-SDWRLS-K---SSW---*ESR-SAVLQA*--L--W--T-S-HPEQ-RN-*VV--EASKDN*QP-VVSTVNTEQ- 33%
Рис. 12. Сравнение аминокислотных последовательностей V (А) и С (Б) областей L-цепи ИГ стерляди с последовательностями других видов позвоночных. Дефис обозначает идентичность в данной позиции. Звездочка обозначает делецию аминокислотных остатков. Слева указаны проценты сходства. Видовые обозначения: Aru - стерлядь, Ipu - пещерный сомик, Omy - радужная форель, Xla - шпорцевая лагушка, Gci - акула-нянька, Hu - человек, Cpl - песчаная акула, Hfr - разнозубая акула.
Рис. 13. Саузерн блот гибридизация. Геномная ДНК была выделена из печени и обработана эндонуклеазами рестрикции Pst I Pvu II. Гибридизацию проводили в мягких условиях с использованием зондов, гомологичных VL (а) и CL (б) генным сегментам L-цепей ИГ стерляди. С V-зондом гибридизовалось более 20 фрагментов ДНК, в то время как с С-зондом гибридизовалось только 3-4 фрагмента.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В результате проведенной работы идентифицирован локус генов, кодирующих L-цепи ИГ стерляди, представителя подкласса костно-хрящевых рыб. Сравнительный анализ структуры V генов этого локуса указывает, что он относится к каппа типу. Результаты сравнительного анализа С областей менее информативны, что само по себе не удивительно, так как С-гены L-цепей ИГ значительно менее консервативны чем V-гены. Тем не менее, тот факт, что на нуклеотидном уровне С-ген стерляди имеет наибольшее сходство с каппа-подобными генами III класса хрящевых рыб, также свидетельствует о том, что обнаруженный локус относится к каппа типу.
кДНК клонов В1, С2, Е4, а также фрагмент, полученный с помощью ПЦР, содержат близкородственные V-гены. Различия между ними сконцентрированы в основном в области 3-го гипервариабельного района. Последовательности двух обнаруженных J-сегментов отличаются по 10 нуклеотидам, т.е., они представляют собой различные геномные сегменты. Незначительные различия обнаружены также между тремя последовательностями С-генов. С-сегмент клона В1 отличается от полного С-сегмента клона С2 по 9 позициям, из которых 5 ведут к замене аминокислоты. По-видимому, эти последовательсти также представляют собой разные зародышевые гены.
Анализ структурной организации локуса с помощью Саузерн блот-гибридизации указывает, что у стерляди имеются множественные VL гены. Точная оценка их количества невозможна, но наличие более 20 полос гибридизации с разной интенсивностью сигнала, по аналогии с многочисленными литературными данными, позволяет говорить о нескольких десятках копий.
При использовании С-зонда количество полос в мягких условиях гибридизации не превышает 4-х. Учитывая возможность аллелизма можно говорить о наличии в этом локусе 2-4х генов.
Ярко выраженное количественное превосходство V генов над С-генами является характерным признаком сегментарной формы организации локуса. У хрящевых и костистых рыб с кластерной организацией генов L-цепей количество V и С генов очень велико и приблизительно одинаково, что отражается в сходной картине блот-гибридизации. Полученные нами результаты являются серьезным основанием для того, чтобы утверждать, что у стерляди организация генов L-цепей каппа-подобного типа имеет сегментарный характер, подобный организации каппа локуса млекопитающих.
Эти данные указывают на то, что переход от кластерной к сегментарной организации произошел в филогенезе позвоночных очень рано, возможно уже у древних первично-костных.
ВЫВОДЫ
1. Сконструирована библиотека кДНК лейкоцитов стерляди представительностью 6х106 независимых клонов. ПЦР ДНК библиотеки с помощью вырожденного праймера, соотвествующего J сегменту генов ИГ, позволил выявить фрагмент ДНК длиной 370 пн, гомологичный V генам ИГ позвоночных.
2. Из библиотеки кДНК с использованием продукта ПЦР в качестве зонда выделено пять клонов, содержащих вставки кДНК генов L-цепей ИГ. Сравнительный анализ первичной структуры кДНК клонов показал, что они являются членами одного семейства генов и имеют наибольшее сходство с генами каппа типа.
3. Согласно результам геномного блот-анализа идентифицированное семейство содержит несколько десятков V генов и только 2-4 С гена, что указывает на сегментарную форму его организации.
4. Полученные данные указывают что переход от кластерного типа организации генов ИГ, характерного для хрящевых рыб, к сегментарному, присутствующему у всех млекопитающих, произошел в эволюции в период появления первично-костных рыб.
СПИСОК ЛИТЕРАТУРЫ
Мазин А. В., Кузнеделов К. Д. и др. Методы молекулярной генетики и генной инженерии. Новосибирск, 1988. 333 с.
Маниатис Т., Фрич Э., Сэмбрук Д. Молекулярное клонирование. М.: Мир, 1984. 480 с.
Пол У. (ред.) Иммунология. М.: Мир, 1987. Т. 1. 476 с.
Ромер А. С. и Парсонс Т. С. Анатомия позвоночных. М.: Мир, 1992. 358 с.
Фримель Г. (ред.) Иммунологические методы. М.: Медицина, 1987. 472 с.
Юдкин И. И. Ихтиология. Москва-Ленинград: Пищепромиздат, 1941.
Aguilera R. J., Akira S., Okazaki K.,Sakano H. A pre-B cell nuclear protein wich specifically interacts with the immunoglobulin V-J recombination sequeces // Cell. 1987. V. 51. P. 909-917.
Anderson M. K., Shamblott M. J., Litman R. T., Litman G. W. Generation of immunoglobulin light chain gene diversity in Raja erinacea is not associated with somatic rearrangement, an exception to a central paradigm of B cell immunity // Journal of Experimental Medecine. 1995. V. 182. P. 109-119.
Bengten E. The immunoglobulin genes in Atlantic cod (Gadus morhuna) and rainbow trout (Oncorhynchus mykiss) // Dissertations from the Faculty of Science and Technology. 1994. V. 19.
Bengten E., Leanderson T., Pilstrom L. Immunoglobulin heavy chain cDNA from Atlantic cod (Gadus morhuna L.): nucleotide sequences of secretory and membrane form show an unusual splicing pattern // European Journal of Immunology. 1991. V. 21. P. 3027-3033.
Blomberg B., Tonegawa S. DNA sequences of the joining regions of mouse l light chain immunoglobulin genes // Proc. Natl. Acad. Sci. USA. 1982. V. 79. P. 530.
Boyle J. S., Lew A. M. An inexpensive alternative to glassmilk for DNA purification // Trends in Genetics. 1995. V. 11. No. 1. p. 8.
Chomczynski P. and Sacchi N. Single-step method of RNA isolation by acid guanidinum thiocyanate-phenol-chloroform extraction // Analytical biochemistry. 1987. V. 162. P. 156-159.
Daggfeldt A., Bengten E., Pilstrom L. A claster type organization of the loci of the immunoglobulin light chain in Atlantic cod (Gadus morhuna L.) and rainbow trout (Oncorhynchus mykiss Walbaum) indicated by nucleotide sequences of cDNAs and hybridization analysis // Immunogenetics. 1993. V.38. P. 199-209.
Dieffenbach C. W. and Dveksler G. S. (edit). PCR primer. A laboratory manual. New York: Cold Spring Harbor Laboratory Press, 1995. 714 p.
Durdik J., Moore M. W., Selsing E. Novel k light-chain gene rearrangements in mouse l light-chain-producing B limphocytes // Nature. 1984. V. 307. P. 749-752.
Flajnik M. F. The immune system of ectothermic vertebrates // Veterinary Immunology and immunopathology. 1996. V. 54. P. 145-150.
Ghaffari S. H. and Lobb C. J. Structure and genomic organization of immunoglobulin light chain in the channel catfish // The Journal of Immunology. 1993. V. 151. P. 6900-6912.
Gilbert W. Evolution of antibodies: the road not taken // Nature. 1990. V. 320. P. 485.
Greenberg A. S., Steiner L.,Kasahara M., Flajnik M. F. Isolation of a immunoglobulin light chain cDNA clone encoding a protein resembling mammalian k light chains: Implications for the evolution of light chains // Proc. Natl. Acad. Sci. USA. 1993. V. 90. P. 10603-10607.
Haire R. N., Ota T., Rast J. P., Litman R. T., Chan F. Y., Zon L. I., Litman G. W. A third Ig light chain gene isotype in Xenopus laevis consists of six distinct VL families and is related to mammalian l genes // The Journal of Immunology. 1996. V. 157. P. 1544-1550.
Hesse J. E., Lieber M. R., Mizuuchi K., Gellert M. V(D)J recombination: the functional difinition of the joining signals // Genes Dev. 1989. V. 3. P. 1053-1061.
Hieter P. A., Hollis G. F., Korsmeyer S. J., Waldmann T. A., Leder P. Clastered arrangement of immunoglobulin l constant region genes in man // Nature. 1981. V. 294. P. 536.
Hieter P. A., Maizel J. V., Leder J., Leder P. Evolution of human immunoglobulin k J region genes // J. Biol. Chem. 1982. V. 257. P. 1516.
Hohman V. S., Schluter S.F., Marchalonis J. J. Complete sequence of a cDNA clone specifying sandbar shark immunoglobulin light chain: Gene organization and implications for the evolution of light chains // Proc. Natl. Acad. Sci. USA. 1992. V. 89. P. 276-280.
Hohman V. S., Schuchman D. B., Schluter S.F., Marchalonis J. J. Genomic clone for sandbar shark l light chain: Generation diversity in the absence of rearrangement // Proc. Natl. Acad. Sci. USA. 1993. V. 90. P. 9882-9886.
Hsu E., Lefkovits I., Flajnik M., Pasquier L. D. Light chain heterogeneity in the amphibian Xenopus // Molecular Immunology. 1991. V. 28. P. 985-994.
immunoglobulin genes and the antibody repertoire // Molecular Biology Evolution. 1993. V. 10(1). P. 60-72.
Litman G. W., Rast J. p., Shamblott M. J., Haire R. N., Michele H., Roess W., Litman R. T, Hinds-Frey K. R., Zilch A., Amemiya C. T. Phylogenetic diversification of Lobb C. J., Olson M. O. J., Clem W. L. Immunoglobulin light chain classes in a teleost fish // The Journal of Immunology. 1984. V. 132. P. 1917-1923.
Lundqvist M., Bengten E., Stromberg S., Pilstrom L. Ig light chain gene in the siberiah sturgeon (Acipenser baeri) // The Journal of Immunology. 1996. V. 157. P. 2031-2038.
Maisey J. G. Chondrichthyan phylogeny: a look at the evidence // J. Ven. Paleont. 1984. V. 4. P. 359-371.
Rast J. P., Anderson M. k., Ota T. Litman R. T., Margittai M., Shamblott M. J., Litman G. W. Immunoglobulin light chain class multiplisity and alternative organizational form in early vertebrate philogeny // Immunogenetics. 1994. V. 40. P. 83-89.
Reynaund C. A., Anquenz V., Grimal H., Weill J. C. A hyperconversion mechanism generates the chicken light chain preimmune repertoire // Cell. 1987. V. 48. P. 379-388.
Roitt I., Brodstoff J., Male D. Immunology. London: Mosby-Year Book Europe, 1993.
Romer A. S. Vertebrate Paleontology. Chicago: University Chicago Press, 1966.
Sakano H., Huppi K., Heinrich G., Tonegawa S. Sequences at the somatic recombination sites of immunoglobulin light-chain genes // Nature. 1979. V. 280. P. 288.
Sakano H., Maki R., Kurosawa Y., Roeder W., Tonegawa S. Two types of somatic recombination are necessary for the generation of complete immunoglobulin heavy-chain genes // Nature. 1980. V. 286. P. 676.
Sanger F., Niklen, S., Coulson A.R. DNA sequencing with chain-terminating inhibitors // Proc. Natl. Acad. Sci. U S A. 1977. V. 74. P. 5463-5467
Schatz G. D., Oettinger A. M., Schlissel M. S. V(D)J recombination: molecular biology and regulation // Annual Review of Immunology. 1992. V. 10. P. 359-383.
Schwager J., Burckert N., Schwager M., Wilson M. Evolution of immunoglobulin light chain genes: analysis of Xenopus IgL isotypes and their contribution to antibody diversity // The EMBO Journal. 1991. V. 10. P. 505-511.
Seidman J. G., Max E. E., Leder P. A. A k immunoglobulin gene is formed by site - specific recombination without further somatic mutation // Nature. 1979. V. 280. P. 370.
Shamblott M. J. and Litman G. W. Genomic organization and sequences of immunoglobulin light chain genes in a primitive vertebrate suggest coevolution of immunoglobulin gene organization // The EMBO Journal. 1989. V. 8. P. 3733-3739.
Shankey T.V. and Clem L.W. Phylogeny of immunoglobulin structure and function. IX. Intramolecular heterogeneity
of shark 19S IgM antibodies to the dinitrophenyl hapten //
J. Immunol. 1980. V. 125. P. 2690-2698.
Stavnezer J., Kekish O., Batter D., Grenier J., Balazs I., Henderson E., Zegers B. J. Aberrant recombination events in B cell lines derived from k-deficient human // Nucl. Acids Res. 1985. V. 13. P. 3495-3514.
Stewart S. E., Pasquier L. D., Steiner L. A. Diversity of expressed V and J regions of immunoglobulin light chains in Xenopus laevis // European Journal of Immunology. 1993. V. 23. P. 1980-1986.
Strob U. Transgenic mise with immunoglobulin genes // Annual Review of Immunology. 1987. V. 5. P. 151-174.
Thompson G. B. and Neiman P. E. Somatic diversification of the chicken immunoglobulin light chain gene is limited to the rearranged variable gene segment //Cell. 1987. V. 48. P. 369-378.
Tonegawa S. Somatic generation of antibody diversity // Nature. 1983. V. 302. P. 575-581.
Udey J. A., Blomberg B. Human l light chain locus: organization and DNA sequences of three genomic J regions // Immunogenetics. 1987. V. 25. P. 63.
Wai Lin Tung, King-C. Chow. A modified medium for efficient electrotransformation of E.coli // Trends in Genetics. 1995. V. 11. No. 4. Р. 128-129.
Wilson M., Hsu E., Marcuz A., Courent M., Pasquier L. D., Steinberg C. What limits affinity maturation of antibodies in Xenopus: the rate of somatic mutation or the abilityto select mutants? // The EMBO Journal. 1992. V. 11. P. 4337.
Zezza D. J., Mikoryak C. A., Schwager J., Steiner L. A. Sequence of C region of L chains from Xenopus laevis Ig // The Journal of Immunology. 1991. V. 146. P. 4041-4047.
Zezza D. J., Stewart S. E., Steiner L. A. Genes encoding Xenopus laevis Ig L chains. Implications for the evolution of k and l chains // The Journal of Immunology. 1992. V. 149. P. 3968-3977.
МИНИСТЕРСТВО ОБЩЕГО И ПРОФЕССИОНАЛЬНОГО
ОБРАЗОВАНИЯ РФ
НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ФАКУЛЬТЕТ ЕСТЕСТВЕННЫХ НАУК
Кафедра цитологии и генетики
КЛОНИРОВАНИЕ И АНАЛИЗ ГЕНОВ ЛЕГКИХ ЦЕПЕЙ ИММУНОГЛОБУЛИНОВ СТЕРЛЯДИ
Дипломная работа
Денисова С. Г.
_____________
Научный руководитель
к.б.н. Таранин А. В.
_____________
Работа выполнена в
лаборатории иммуногенетики
Института цитологии и
генетики СО РАН.
Новосибирск 1997