Реферат: Модели нейронных сетей
1. Нейроподобный элемент (нейрон).
На нейроподобный элемент поступает набор входных сигналов x1, x2, ..., xM (или входной вектор X), представляющий собой выходные сигналы других нейроподобных элементов. Каждый входной сигнал умножается на соответствующий вес связи w1, w2, ..., wM - аналог эффективности синапса. Вес связи является скалярной величиной, положительной для возбуждающих и отрицательной для тормозящих связей. Взвешенные весами связей входные сигналы поступают на блок суммации, соответствующий телу клетки, где осуществляется их алгебраическая суммация и определяется уровень возбуждения нейроподобного элемента S:
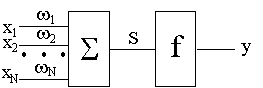
Рис. 1.1.
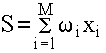 (1.1)
(1.1)
Выходной сигнал нейрона y определяется путем пропускания уровня возбуждения S через нелинейную функцию f:
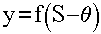 (1.2)
(1.2)
где q - некоторое постоянное смещение (аналог порога нейрона). Обычно используются простейшие нелинейные функции: бинарная (рис. 1.2, а)
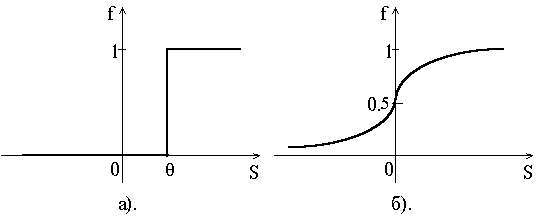
Рис. 1.2.
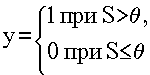 (1.3)
(1.3)
или сигмоидная (рис. 1.2, б)
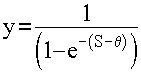 (1.4)
(1.4)
В такой модели нейрона пренебрегают многими известными характеристиками биологического прототипа, которые некоторые исследователи считают критическими. Например, в ней не учитывают нелинейность пространственно-временной суммации, которая особенно проявляется для сигналов, приходящих по возбуждающим и тормозящим синапсам, различного рода временные задержки, эффекты синхронизации и частотной модуляции, рефрактерность и т.п. Несмотря на это нейроподобные сети, построенные на основе таких простых нейроподобных элементов, демонстрируют ассоциативные свойств, напоминающие свойства биологических систем.
2. Нейроподобная сеть.
Нейроподобная сеть представляет собой совокупность нейроподобных элементов, определенным образом соединенных друг с другом и с внешней средой. Входной вектор (координирующий входное воздействие или образ внешней среды) подается на сеть путем активации входных нейроподобных элементов. Множество выходных сигналов нейронов сети y1, y2, ...,yN называют вектором выходной активности, или паттерном активности нейронной сети. Веса связей нейронов сети удобно представлять в виде матрицы W, где w ij - вес связи между i- и j-м нейронами. В процессе функционирования (эволюции состояния) сети осуществляется преобразование входного вектора в выходной, т.е. некоторая переработка информации, которую можно интерпретировать, например, как функцию гетеро- или автоассоциативной памяти. Конкретный вид выполняемого сетью преобразования информации обусловливается не только характеристиками нейроподобных элементов, но и особенностями ее архитектуры, т.е. той или иной топологией межнейронных связей, выбором определенных подмножеств нейроподобных элементов для ввода и вывода информации или отсутствием конкуренции, направлением и способами управления и синхронизации информационных потоков между нейронами и т.д.
2.1. Модели нейронных сетей.
Рассматриваемые нами модели нейронных сетей объединены в три группы. В п. 2.1.1. рассматриваются сети персептронного типа, для которых характерно отсутствие обратных связей между нейроподобными элементами, организованными в слои. Отличительной особенностью сетей, представленных в п. 2.1.2, являются симметричные (равные по величине и противоположные по направлению) связи между любыми двумя соединенными нейронами. В нейросетевых архитектурах, описанных в п. 2.1.3, между нейронами одного слоя имеются постоянные тормозящие связи (латеральное торможение).
2.1.1. Сети с прямыми связями.
Прямой персептрон. В середине 50-х годов была предложена одна из первых моделей нейронных сетей, которая вызвала большой интерес из-за своей способности обучаться распознаванию простых образов. Эта модель - персептрон - состоит из бинарных нейроподобных элементов и имеет простую топологию, что позволило достаточно полно проанализировать ее работу и создать многочисленные физические реализации. Типичный персептрон состоит из трех основных компонент:
матрицы бинарных входов r1, r2, ..., rn (сенсорных нейронов или "сетчатки", куда подаются входные образы);
набора бинарных нейроподобных элементов x1, x2, ..., xm (или предикатов в наиболее общем случае) с фиксированными связями к подмножествам сетчатки ("детекторы признаков");
бинарного нейроподобного элемента с модифицируемыми связями к этим предикатам ("решающий элемент").
На самом деле число решающих элементов выбирают равным количеству классов, на которое необходимо разбить предъявляемые персептрону образы.
Таким образом, модель персептрона характеризуется наличием только прямых связей, один из слоев которых является модифицируемым. В постейшем случае, когда n = m и xi = ri, детекторы признаков могут рассматриваться как входной слой. Тогда персептрон становится одним бинарным нейроподобным элементом. Это классическая модель М-входового нейрона, приведенная на рис. 1.1, или простой персептрон Розенблатта. В общем случае каждый элемент xi может рассматриваться как булева функция, зависящая от некоторого подмножества сетчатки. Тогда величина выходных сигналов этих обрабатывающих элементов является значением функции xi, которое равно 0 или 1.
Устройство реагирует на входной вектор генерацией выходного сигнала y решающего элемента по формуле (1.3). Таким образом, персептрон формирует гиперплоскость, которая делит многомерное пространство x1, x2, ..., xm на две части и определяет, в какой из них находится входной образ, выполняя таким образом, его классификацию. Возникает вопрос, как определить значения весов, чтобы обеспечить решение персептроном конкретной задачи. Это достигается в процессе обучения.
Один из алгоритмов обучения приведен в параграфе 2.2.
Многослойный персептрон. Как отмечалось выше, простой персептрон с одним слоем обучаемых связей формирует границы областей решений в виде гиперплоскотей. Двухслойный персептрон может выполнять может выполнять операцию логического "И" над полупространствами, образованными гиперплоскостями первого слоя весов. Это позволяет формировать любые, возможно неограниченные, выпуклые области в пространстве входных сигналов. С помощью трехслойного персептрона, комбинируя логическими "ИЛИ" нужные выпуклые области, можно получить уже области решений произвольной формы и сложности, в том числе невыпуклые и несвязные. То, что многослойные персептроны с достаточным множеством внутренних нейроподобных элементов и соответствующей матрицей связе в принципе способны осуществлять любое отображение вход - выход, отмечали еще Минский и Пейперт, однако они сомневались в том, что можно открыть для них мощный аналог процедуры обучения простого персептрона. В настоящее время в результате возрождения интереса к многослойным сетям предложено несколько таких процедур. Часть из них приведена в параграфе 2.2.
2.1.2. Сети с симметричными связями.
Ансамблевые нейронные сети. Минский и Пейперт отмечали, что недостатки простых персептронов можно преодолеть как с помощью многослойных сетей (см. выше), так и введением в сеть обратных связей, допускающих циркуляцию сигналов по замкнутым контурам. Использовать свойства такого рода сетей для моделирования функций мозга еще в 1949 г. предложил Хебб.
Согласно взглядам Хебба нервные клетки мозга соединены друг с другом большим количеством прямых и обратных возбуждающих связей и образуют нейронную сеть. Каждый нейрон осуществляет пространственно-временную суммацию приходящих к нему сигналов от возбуждающих, определяя потенциал на своей мембране. Когда потенциал на мембране превышает пороговое значение, нейрон возбуждается. Нейрон обладает рефрактерностью и усталостью. Эффективность связей может изменяться в процессе функционирования сети, повышаясь между одновременно возбужденными нейронами. Это приводит к объединению нейронов в клеточные ансамбли - группы клеток, которые чаще всего возбуждались вместе, и к обособлению ансамблей друг от друга. При возбуждении достаточной части ансамбля он возбуждается целиком. Различные ансамбли могут пересекаться: один и тот же нейрон может входить в разные ансамбли. Электрическая активность мозга обусловлена последовательным возбуждением отдельных ансамблей.
Идеи Хебба оказали большое воздействие на представления о работе мозга и послужили основой для создания нейронных моделей долговременной памяти. Действительно, ансамблевую нейронную сеть можно рассматривать как структуру, реализующую функции распределенной ассоциативной памяти. Формирование ансамблей в такой сети соответствует запоминанию образов (признаков, объектов, событий, понятий), закодированных паттерном активности нейронов, а сформированные ансамбли являются их внутренним представлением. Процесс возбуждения всего ансамбля при активации части его нейронов можно интерпретировать как извлечение запомненной информации по ее части - ключу памяти.
Модель памяти на основе ансамблевой нейронной сети обладает некоторыми свойствами, присущими биологической памяти, таким, как ассоциативность, распределенность, параллельность, устойчивость к шуму и сбоям, надежность. Проводятся также структурные аналоги между ансамблевыми моделями нейронных сетей и строением коры головного мозга. Имеются экспериментальные данные о синаптической пластичности, постулированной Хеббом.
Модель ансамблиевой сети состоит из большого количества нейронов, каждый из которых обычно соединен со всеми другими элементами сети. Входной образ подается на сеть путем активации нужных нейроподобных элементов. Обучение ансамблиевой сети описано в параграфе .
Сеть Хопфилда. Хотя многочисленные результаты моделирования демонстрировали стабильность ансамблевых сетей с обратными связями и хеббовским правилом обучения (эволюцию сети к устойчивому состоянию), отсутствие математического обоснавания такого поведения препятствовало их популярности.
В 1982 г. американский биофизик Джон Хопвилд опубликовал статью, где поведение модели полносвязной сети бинарных нейроподобных элементов с симметричными связями (w ij = w ji). элементы функционировали в асинхронном режиме, т.е. каждый нейрон в случайные моменты времени с некоторой средней частотой определял свое состояние с правилом (1.3). Это позволило описать поведение сети ка релаксационный процесс, при котором минимизируется функция (гамильтониан) модели.
Машина Больцмана представляет собой стохастический вариант сети Хопфилда. Бинарные нейроподобные элементы (блоки) трактуются здесь представители элементарных гипотез, а веса - как слабые парные взаимоограничения между ними. Положительный вес связи указывает, что две гипотезы стремятся поддерживать друг друга, а отрицательный - на их несовместимость. Симметрия связей позволяет проанализировать поведение сети с использованием энергетической функции (гамильтониана). Энергию определенного паттерна активности можно интерпретировать как степень нарушения ограничений, присутствующих в проблемной области, со стороны конкретной комбинации гипотез или как стоимостную функцию, которая должна быть минимизирована для решения оптимизационной задачи. Существует возможность, однако, попадания сети в локальный мимнимум, что крайне не желательно для оптимизационных задач. Чтобы сеть могла выбраться из локального энергетического минимума, в машине Больцмана применяется вероятностное правило срабатывания блоков:
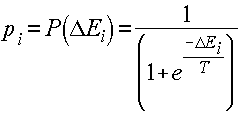 , (2.1)
, (2.1)
где pi - вероятность нахождения i-го блока в единичном состоянии;
P (x) - сигмоидная функция (рис. 1.2. б);
T - параметр, аналогичный температуре. При T® 0 это правило переходит в правило срабатывания детерминированных элементов (1.3), а при повышении температуры увеличивается вероятность перехода системы в состояние с большей энергией.
Обучение машины Больцмана описано в параграфе 2.2.
2.1.3. Сети с латеральным торможением.
Карты признаков Кохонена. Обычно в качестве входных образов в моделях ассоциативной памяти используются некоторые внутренние представления сенсорной информации, прошедшей, как считается, необходимую предобработку. Один из нейросетевых вариантов такой переработки предложен Кохоненом. Его алгоритм формирует одно- или двумерную карту "карту" признаков путем нелинейного "сплющивания" многомерного сигнального пространства. При этом предполагается, что такое отображение должно сохранять топологические отношения, существующие между входными сигналами.
Структура нейронной сети, в которой реализуется формирование карт признаков, приведена на рис. 2.1. Нейроны, имеющие сигмоидную характеристику, расположены в виде одно- и двумерного слоя слоя по аналогии со слоистым строением коры. На каждый нейрон поступают два вида связей: mij, которые интерпретируются как связи от сенсорных входов или из других областей, и w jk - латеральные связи от нейронов одного слоя, характер которых зависит от расстояния между нейронами. Функция взаимодействия нейронов одного слоя имеет вид "мексиканской шляпы" (рис. 2.2.), что соответствует некоторым нейробиологическим данным. Близко расположенные нейроны возбуждают друг друга, с увеличением расстояния возбуждение сменяется торможением, а затем опять появляются слабые возбуждающие связи, которые по-видимому, выполняют ассоциативные функции и в данной модели не используются.
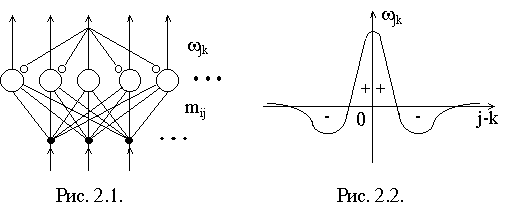
Эффект наличия латеральных связей с радиусом действия порядка размеров сети проявляется в следующем. Если на каждый нейрон подать (например, через связи от сенсорных входов mij) имеющий небольшой максимум случайный сигнал Si, то в процессе релаксации сети осуществляется повышение его контрасности. В результате вокруг первоначального максимума образуется "пузырек" выходной активности нейронов (рис. 2.3.).
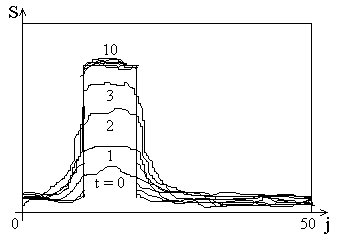
Рис. 2.3.
Входные сигналы полностью определяют процесс самоорганизации сети, т.е. в ней реализован алгоритм обучения без учителя. Латеральные связи w jk в модели считаются постоянными, и все адаптивные эффекты происходят только в матрице входных связей М. Подробнее процесс обучения рассмотрен в параграфе 2.2.
Теория адаптивного резонанса. Пожалуй, одна из самых развитых и продуманных с биологической точки зрения концепций нейросетевой обработки информации предложена в работах Гроссберга. Ее стержнем является модель нейронной сети и алгоритмы теории адаптивного резонанса, которая была разработана в начале 70-х годов и детализирована в 80-х.
Нейронная система теории адаптивного резонанса способна обучаться распознаванию образов различной степени сложности. Она относит входной образ к одному из классов в зависимости от того, на какой образ из запомненных образов он больше всего похож. Если входной образ не соответствует ни одному из запомненных, создается новый класс путем его запоминания. Если найден образ, с определенным "допуском" соответствующий входному, то он модифицируется так, чтобы стать еще больше похожим на входной.
2.2. Обучение нейронной сети.
Одно из важнейших свойств нейроподобной сети - способность к самоорганизации, самоадаптации с целью улучшения качества функционирования. Это достигается обучением сети, алгоритм которого задается набором обучающих правил. Обучающие правила определяют, каким образом изменяются связи в ответ на входное воздействие. Обучение основано на увеличении силы связи (веса синопса) между одновременно активными нейронами. Таким образом, часто используемые связи усиливаются, что объясняет феномен обучения путем повторения и привыкания. Математически это правило можно записать так:
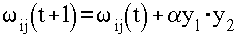 (1.5)
(1.5)
где w ij(t) и w ij(t+1) - значение связи от i-го к j-му нейрону соответственно до и после его изменения, a - скорость обучения. В настоящее время существует множество разнообразных обучающих правил (алгоритмов обучения). Некоторые из них приведены ниже.
2.2.1. "Back propagation" (алгоритм обратного распространения ошибки).
Этот алгоритм является обобщением одной из процедур обучения простого персептрона, известной как правило Уидроу - Хоффа (или дельта-правило), и требует представления обучающей выборки. Выборка состоит из набора пар образов, между которыми надо установить соответствие, и может рассматриваться как обширное задание векторной функции, область определения которой - набор входных образов, а множество значений - набор выходов.
Перед началом обучения связям присваиваются небольшие случайные значения. Каждая итерация процедуры состоит из двух фаз. Во время первой фазы на сеть подается входной вектор (образ) путем установки в нужное состояние входных элементов. Затем входные сигналы распространяются по сети, порождая некоторый выходной вектор. Для работы алгоритма требуется, чтобы характеристика вход - выход нейроподобных элементов была неубывающей и имела ограниченную производную. Обычно для этого используют сигмоидную нелинейность вида (1.4).
Полученный выходной вектор сравнивается с требуемым. Если они совпадают, обучения не происходит. В противном случае вычисляется разница между фактическими и требуемыми выходными значениями, которая передается последовательно от выходного слоя к входному. На основании этой информации об ошибке производится модификация связей с обобщенным дельта-правилом.
2.2.2. Обучение без "воспитателя".
Обучение без "воспитателя" возможно например в сетях адаптивного резонанса (параграф 2.1.3.). Происходит сравнение входного образа с имеющимися в памяти сети шаблонами. Если нет подходящего шаблона, с которым можно было бы отождествить исследуемый образ, то создается новый шаблон, содержащий в себе этот входной образ. В дальнейшем новый шаблон используется наравне с другими.
3. Нейрокомпьютеры.
Термин "нейрокомпьютер" употребляется для обозначения всего спектра работ в рамках подхода к построению систем искусственного интеллекта, основанного на моделировании элементов, структур, взаимодействий и функций различных нервной системы. Так как в настоящее время исследования в этой области ведутся в основном на уровне моделей нейронных сетей, то понимание термина "нейрокомпьютеры" сужают, ставя знак равенства между ним и нейронными сетями.
В зависимости от способа реализации моделей нейронных сетей выделяют 4 уровня нейрокомпьютеров.
Теоретический. Работы, в которых в той или иной форме (математической, алгоритмической, словесной и т.д.) представлено описание моделей нейронных сетей.
Программный. Модели нейронных сетей, программно реализованные на обычных последовательных компьютерах.
Программно-аппаратный. Сопроцессоры для ускорения моделирования нейронных сетей.
Аппаратный. Физически реализованные модели нейронных сетей.
Специфичность нейросетевых операций, а также сверхпараллельность структуры и функционирования моделей нейронных сетей чрезвычайно замедляют их реализацию на обычных последовательных компьютерах. Потребность в выполнении большого объема исследовательских работ и быстром функционировании появившихся прикладных систем привели к появлению специализированных вычислительных устройств для эффективного моделирования нейронных сетей - нейрокомпьютеров в узком смысле слова. Такая трактовка, соответствующая уровням 2 и 3 по приведенной классификации, получила широкое распространение.
Заключение.
Рассмотренные нами нейроподобные сети могут выполнять большой круг задач.
| Технология извлечения знаний из нейронных сетей: апробация ... | |
|
... Омский Государственный Университет Математический факультет Горбань Павел Александрович ТЕХНОЛОГИЯ ИЗВЛЕЧЕНИЯ ЗНАНИЙ ИЗ НЕЙРОННЫХ СЕТЕЙ: АПРОБАЦИЯ, ... Для полносвязной сети входной сумматор нейрона фактически распадается на два: первый вычисляет линейную функцию от входных сигналов сети, второй - линейную функцию от выходных ... Рассматривалась ситуация активации/неактивации нейрона при конкретных комбинациях значений бинарных входных сигналов нейрона. |
Раздел: Рефераты по информатике, программированию Тип: дипломная работа |
| Психологическая интуиция искусственных нейронных сетей | |
|
министерство общего и специального образования Российской Федерации сибирский государственный технологический университет На правах рукописи Доррер ... где mi0 - параметр инциации сети, xi1 - входные сигналы сети, поступающие на данный нейрон, fiT - выходной сигнал нейрона на такте времени T, Ai1 - входной параметр i-го нейрона на ... Такая информация полезна в тех случаях, когда размерность вектора входных сигналов велика и время обучения также оставляет желать лучшего, поскольку позволяет уменьшить размерность ... |
Раздел: Рефераты по информатике, программированию Тип: дипломная работа |
| Классификация сейсмических сигналов на основе нейросетевых технологий | |
|
... МАТЕМАТИКИ Кафедра МОСОИиУ ПОЯСНИТЕЛЬНАЯ ЗАПИСКА К ДИПЛОМНОМУ ПРОЕКТУ На тему: _Классификация сейсмических сигналов на основе нейросетевых технологий. В теории нейронных сетей разработаны более мощные алгоритмы, осуществляющие "нелинейный анализ главных компонент"[3]. Они представляют собой самостоятельную нейросетевую структуру ... Автоматически, в зависимости от размерности входных векторов, определяется конфигурация нейронной сети т.е. по умолчанию для заданной размерности входных данных выбирается ... |
Раздел: Рефераты по кибернетике Тип: реферат |
| Системное автоматизированное проектирование | |
|
Радиотехники, Электроники и Автоматики Московский Государственный Институт(Технический Университет) С.А.Шишов Лекции по дисциплине: "Системное ... Если нет соответствия между входным и выходным векторами, то веса связей, соединяющих активные входные и выходные нейроны, уменьшаются на ту же величину del. Выходной слой персептрона пусть содержит один бинарный нейрон. |
Раздел: Рефераты по информатике, программированию Тип: реферат |
| Математические модели в естествознании | |
|
Вопросы экзамена 1. Основные понятия и определения генетики. 2. Законы Менделя 3. Закон Харди- Вайнберга 4. Принцип стационарности. Кадрильный закон ... Если входной вектор , то выходной сигнал нейрона , если же , то . Требуется выбрать синаптическую матрицу и пороговый вектор так, чтобы для входных векторов персептрон формировал выходные векторы , т.е. |
Раздел: Рефераты по математике Тип: шпаргалка |