Стратиграфическая и геохронологическая таблица Триасового периода Мезозойской эры
Министерство Образования и Науки Российской Федерации
Государственное Образовательное Учреждение
Оренбургский Государственный Университет.
Кафедра Геологии
Факультет Вечернего и Заочного Обучения
Курсовая на тему: а)Стратиграфическая и геохронологическая таблица Триасового периода Мезозойской эры ; б)Характеристика типа Артропода(Членистоногие).
Выполнил: студент Вечернего и Заочного обучения
Курс 1 Группа 07 ГС Специальность ГС
Проверил:
План.
1)Стратиграфическая и геохронологическая таблица Триасового периода Мезозойской эры.
2)Характеристика типа Arthropoda (Членистоногие).
3) Список использованной литературы.
Введение.
В ряду естественных наук почётное место занимает - Палеонтология. В буквальном переводе палеонтология означает – знание, наука о древней жизни. Отсюда ясно, что палеонтология изучает представителей животного и растительного мира прошлых геологических эпох. Изучение этих событий немыслимо без стратиграфических и геохронологических исследований. При стратиграфических исследованиях осуществляют две последовательные операции:
1) расчленение разреза на отдельные стратиграфические подразделения (слои, пачки, горизонты, подъярусы, ярусы и т. д.) на основании различий состава горных пород и заключенных в них ископаемых органических остатков, а также проявлений перерывов и несогласий;
2) сопоставление или стратиграфическая корреляция выделенных в разных разрезах слоев, пачек, горизонтов, т. е. установление их геологической одновозрастности.
Фактической основой стратиграфических исследований служат конкретные геологические объекты — естественные или искусственные обнажения горных пород и керн скважин, а также определяемые геофизическими методами (электро-, сейсмо- и другой каротаж) изменения физических свойств горных пород в скважинах.
После обобщения ряда частных геологических разрезов составляется сводная стратиграфическая колонка, в которой все слои горных пород располагаются в строгой последовательности своего образования и залегания, т. е. в определенном хронологическом порядке, обычно от более древних внизу к более молодым вверху.
Геохронология преследует цель восстановить строгую временную последовательность геологических событий, происходивших в прошлом, путем установления хронологических взаимоотношений между накопившимися слоями горных пород, в которых эти события оказались запечатленными.
Палеонтология, как и любая другая наука, изучающая историю развития живой и неживой природы, невозможна без хронологии. Однако хронология — это еще далеко не история, а лишь механическое расположение событий во времени. В прошлом происходило великое множество различных событий, и для того, чтобы ориентироваться среди них, необходимо не только установить формальные временные соотношения между ними, но и найти внутренние связи между событиями и явлениями прошлого и определить их пространственные взаимоотношения и относительное значение. При этом выявляются естественные временные группировки, разграниченные событиями более высоких рангов, и тем самым намечаются последовательные этапы исторического развития. В таком случае речь идет о естественной периодизации геологической истории.
Стратиграфия (лат. stratum — слой, grapho — пишу) — раздел исторической геологии, занимающийся изучением исторической последовательности, первичных взаимоотношений и географического распространения осадочных, вулканогенно-осадочных и метаморфических образований, слагающих земную кору и отражающих естественные этапы развития Земли и населявшего ее органического мира (Геол.словарь. М., 1973).
Перед стратиграфическими исследованиями стоят следующие важнейшие задачи:
1) детальное расчленение разрезов горных пород и выделение разных по рангу стратиграфических подразделений; на их основе создаются местные, региональные и межрегиональные стратиграфические шкалы, которые отражают хронологическую последовательность геологических событий;
2) проведение региональной и межрегиональной стратиграфической корреляции,
3) создание единой стратиграфической и геохронологической шкалы.
Стратиграфия играет важнейшую роль при геологических исследованиях. Без нее немыслимо проводить геологическое картирование, решать проблемы эволюции органического мира, геологического развития отдельных регионов и Земли в целом, реконструировать палеогеографические обстановки. Без детальных стратиграфических исследований невозможно раскрывать сложное строение структур земной коры и проводить поиски и разведку полезных ископаемых.
|
Триасовый |
Поздняя |
Рэтский |
T3r |
225 |
17 |
|
Норийский |
T3n |
5 |
|||
|
Карнийский |
T3k |
230 |
5 |
||
|
Средняя |
Ладинский |
T2l |
235 |
5 |
|
|
Анизийский |
T2a |
240 |
5 |
||
|
Ранняя |
Оленекский |
T1o |
245 |
||
|
Индский |
T1i |
Мезозойская эра. Триасовый период.
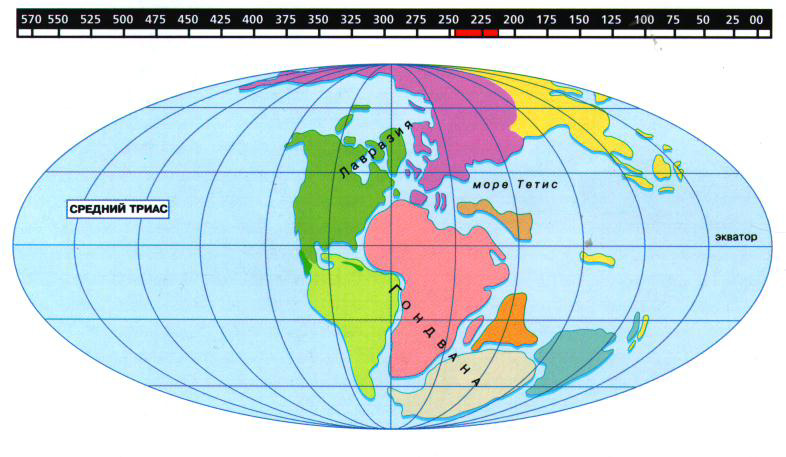
Триасовый период на Земле продолжался около 45 млн. лет. С его начала до наших дней прошло приблизительно 220 млн. лет. В триасе суша преобладала над морем. Существовало два континента. Слившиеся между собой Северо-Атлантический и Азиатский материки образовали Северную сушу. В южном, полушарии лежала прежняя Гондвана. Азия соединилась с Австралией и Новой Зеландией. Вся Южная Европа, Кавказ и Крым, Иран, Гималаи и Северная Африка были залиты океаном Тетке. Больших горных цепей в это время заново не возникало, но горы, сформировавшиеся в прежние периоды, были еще высоки. Часто происходили вулканические извержения. Климат триасового периода был резкий и сухой, но достаточно теплый. Пустыни в триасе многочисленны. Из растений заметно преобладали голосеменные: саговые, хвойные и гинкговые. Из семенных папоротников продолжал существовать глоссоптерис. В конце периода появились своеобразные папоротники, особенно многочисленные в последующем юрском периоде, листочки которых по жилкованию напоминали листья семенных растений. Триасовые хвощи стоят гораздо ближе к современным хвощам, чем палеозойские. В жизни обитателей материков Произошли большие перемены. Начавшиеся еще в пермский период преобладание суши над морем н прогрессивное осушение многих пресных водоемов в триасовый период привели к тому, что многие пресноводные рыбы теперь переселились в моря, и только двоякодышащие, близкие к теперешним, попрежнему жили в уцелевших пресноводных бассейнах. В конце триасового периода вымерли стегоцефалы. Это были последние представители лабиринтозубых стегоцефалов, названных так потому, что эмаль на их зубах имела сложное складчатое строение. Все стегоцефалы, спасаясь от сухого климата и от конкуренции с пресмыкающимися, стали водными, а некоторые даже перешли жить в море. Большинство из них были очень крупными животными. Например, у мастодонтзавра длина черепа достигала 1 м. В начале триасового периода жили прямые предки современных лягушек. Эти протобатрахусы мелкие, длиной в 10 см, животные, по общему складу скорее напоминают жаб, чем настоящих лягушек. Кожа их бугорчатая, задние ноги более приспособлены для плавания, чем для прыжка. Особенно сильно изменились пресмыкающиеся; цельночерепные окончательно вымирали. Во вторую половину периода появились первые черепахи, у которых в отличие от современных на небе еще сохранились зубы, в то время как челюсти были одеты роговым клювом. В триасовый период усиленно развивались, но в конце его уже вымерли последние звероподобные пресмыкающиеся. Из них травоядные и уже совершенно беззубые шталекерии достигали размеров крупного носорога. Меньших размеров был хищный белезодонт длиной около 1,5 м. Особенно интересны мелкие зверообразные пресмыкающиеся иктидозавры, близкие к млекопитающим. Так, каромис, животное величиной с крысу, по строению своего черепа уже является настоящим млекопитающим, и только дополнительные кости, имеющиеся в его нижней челюсти, указывают на то, что это животное еще пресмыкающееся. Из других пресмыкающихся в триасовые период развивались хоботоголовые, ближайшие родичи современной новозеландской гаттерии, которые хотя и похожи на обычных ящериц, но отличаются от них по своему строенною. Гаттерия в своем строении до сих пор сохранила многие древние особенности. В ее черепе имеются две височные (скуловые) дуги, а не одна, как у ящериц. Верхняя челюсть у нее свисает вниз в виде небольшого клюва. Зубы на челюстях сидят не в отдельных ячейках, а в общем желобке. Кроме обычных ребер, на брюхе развиваются еще «брюшные ребра». Двояковогнутые позвонки напоминают позвонки рыб. Среди хоботоголовых в триасе жили стенаулоринхусы – крупные роющие животные, возможно, питавшиеся корнями. В морях, по берегам континентов, попадались длиннорылые хоботоголовые – истребители морских моллюсков. В месте с ними за моллюсками охотились несколько напоминавшие морских черепах плакодонты, у которых на небе вместо мелких зубов образовывались настоящие жерновки для раздавливания раковин. Родственные плакодонтам нотозавры тоже вели водный образ жизни. Эти длинношеие животные еще могли использовать свои лапы (ласты) для хождения по земле. От нотозавров произошли плезиозавры, обычные морские пресмыкающиеся следующих периодов. В северных водах появились первые рыбоящеры, или ихтиозавры. Они еще не так хорошо были приспособлены к плаванию в море, как их потомки, у которых хвост стал похож на рыбий. Самое замечательное то, что ихтиозавры не откладывали яиц, как обычные пресмыкающиеся, а рождали живых детенышей, подобно млекопитающим. С триаса же начался расцвет группы ячеезубых пресмыкающихся. Наиболее древние формы из них были сравнительно небольшие плотоядные животные. Вместо обычного передвижения на четырех ногах эти животные приспосабливались к хождению и на двух, в связи с чем их задние ноги становились гораздо длиннее передних. Таким был сальтопозухус, животное размером больше 1 м. К концу триаса некоторые ячеезубые пресмыкающиеся перешли к водному образу жизни. Они снова начали ходить на четырех ногах и по внешности несколько напоминали еще отсутствовавших тогда крокодилов. Длина такого крокодилообразного престосухуза была не менее 5 м. Преимущественно на Северной суше появились первые, еще не очень большие по размеру редки динозавров. Некоторые из них были не невелики, до 1м в длину, и вели хищный образ жизни. Ходили на задних ногах, которые были длиннее передних. По некоторым признакам динозавры напоминали птиц: кости их скелета были полыми наполненными воздухом, а первый палец на задних нога был обращен назад. Другие динозавры, например платеозавр, были значительно крупнее, достигая 6 м в длину. Разница в строении передних н задних ног у них невелика, зубы тупые. Это были предки травоядных великанов юрского периода. Неудивительно, что при обилии в триасе зверообразных пресмыкающихся мы находим здесь и настоящих млекопитающих. Самое древнее, известное нам млекопитающее животное, величиной с сурка, именуется «тритилодонт». Он относится к группе много бугорчатозубых млекопитающих, называемых так потому, что у них на коренных зубах в два или три ряда были расположены многочисленные бугорки. Клыков у них не было. Одна пара резцов в верхней челюсти и единственная пара в нижней были увеличены. Много бугорчатозубые питались растительной пищей. Вероятно еще откладывали яйца, а не рождали живых Детенышей, так же как и современные австралийские однопроходные млекопитающие: утконос и ехидна. Современные яйцекладущие млекопитающие – беззубы, но зародыши утконоса имеют зачатки зубов многобугорчатого типа. Поэтому много бугорчатозубых считают ближайшими родичами австралийских однопроходных, у которых еще сохраняется много особенностей, характерных для пресмыкающихся. На дне триасового моря жили многочисленные шестилучевые кораллы, близкие к современным. Обильными были двустворчатые и брюхоногие моллюски, которые замещали плеченогих. Часто попадались новые морские ежи и лилии. Но особенного разнообразия в этом периоде достигали многочисленные аммониты. В это же время появились первые белемниты – животные, близкие к современным морским каракатицам, тоже относившиеся к головоногим моллюскам. Под кожей у них был спрятан известковый скелет в виде пластинке, кончавшейся острым шипом. Этот шип обычно сохраняется в ископаемом состоянии и называется «чертов палец».
В море, кроме акуловых рыб, уже жило довольно много костных, предки которых переселились сюда из пресной воды. Здесь встречались кистеперые рыбы и родичи современных, осетровых рыб, а также панцирных щук и ильных рыб Северной Америки. По строению чешуи, хвоста и внутренних органов эти рыбы еще отличались от настоящих костистых рыб.
Триасовый период в истории Земли ознаменовал собой начало мезозойской эры, или "эры средней жизни". До него все материки были слиты в единый гигантский сверхматерик Пангею. С наступлением триаса Пангея начала постепенно раскалываться. Климат в те времена был ровным по всему земному шару. Даже у полюсов и на экваторе погодные условия были гораздо более сходными, чем в наши дни. Ближе к концу триаса климат стал суше. Озера и реки начали быстро пересыхать, и во внутренних областях материков образовались обширные пустыни. Когда в пермский период материки соединились, образовав сверхматерик Пангею, громадные массивы суши наползли друг на друга и поглотили значительную часть побережья Мирового океана. Затем, в триасовый период, климат потеплел, и многие уцелевшие мелководные моря высохли, а оставшаяся вода сделалась очень соленой. Немало прежних форм морской жизни исчезло, и их место заняли новые виды животных.
{kind=link}
Расцвет беспозвоночных. В морях появились новые моллюски, такие, как устрицы. Они зарывались в донный песок, пропускали через свои раковины воду и отфильтровывали из нее частицы пищи. Появилось и множество новых брюхоногих моллюсков (улиток и их родственников). С понижением уровня воды в мелких морях их скалистые берега все более обнажались. Здесь находили себе пристанище новые виды моллюсков, например блюдечки и литорины. Встречались и новые разновидности кораллов, креветок и омаров. В триасе появились также первые "настоящие" морские ежи, похожие на современных. В морях по-прежнему обитали аммониты. В конце триаса они почти все вымерли, а те, кто выжил, "дотянул" до юрского периода, когда наступил их новый расцвет.
{kind=link}
В триасовый период внутри материков было жарко и сухо. Там раскинулись обширные бесплодные пустыни, где было очень мало растений. Однако вблизи побережья попадались плодородные участки земли, богатые всевозможной растительностью. 1 листрозавр 2 ринхозавр 3 гинкго 4 араукария 5 тис 6 саговник 7 древовидный папоротник 8 беннеттит 9 плаун 10 хвощ
От акул до "рыболовных удочек". Дальше от берега в открытом море обитали наиболее высокоорганизованные виды рыб. Акулы и костные рыбы оспаривали друг у друга добычу. Со временем у них развились челюсти, способные разгрызать панцири крабов и раковины моллюсков вроде мидий. Крупнейшими хищниками триасовых морей были недавно возникшие водные рептилии. Ящерицеобразные нотозавры ловили рыбу при помощи своих острых зубов. Дельфиноподобные ихтиозавры могли догнать любую добычу благодаря своим выдающимся скоростным качествам. Крупные плакодонты, похожие на тритонов, ползали по морскому дну, выискивая раковины, а затем раздавливали их мощными плоскими зуоами. У танистрофея была длинная и тонкая шея, вдвое длиннее его туловища. Это было наземное животное, и, вероятно, оно использовало свою изящную шею в качестве рыболовной удочки. Стоя у кромки воды, танистрофей мог дотянуться до рыб, плававших под водой в изрядном удалении от берега.
{kind=link}
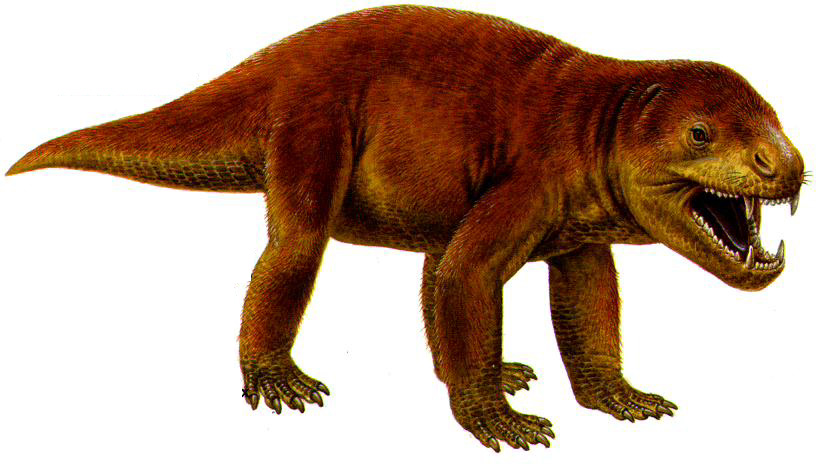
Циногнат, представитель цинодонтов ("собакозубых" рептилий). Это было сильное животное величиной с волка, со многими чертами, свойственными лекопитающим. Ученые не сомневаются, что циногнат имел волосяной покров: при изучении его ископаемых останков у него на морде обнаружили ямки, из которых росли усы. Значит, возможно, что эти животные были теплокровными, ибо волосяной покров до сихпор встречался лишь у теплокровных млекопитающих.
Триасовые стада. В начале триасового периода животный мир был одинаков на всей территории земной суши. Различные виды могли беспрепятственно распространяться по всей Пан-гее, ибо на их пути не возникало больших водных преград. Многие животные, обитавшие на Земле в пермский период, вымерли к началу триаса, возможно, из-за климатических изменений. Но некоторые из зверообразных рептилий все же уцелели, причем иные виды в больших количествах. Громадные стада растительнояд-ных листрозавров нежились в иле по берегам озер и рек. Это были настоящие "бегемоты" триасового мира. Их ископаемые останки нашли в таких удаленных друг от друга местностях, как Китай, Индия, Южная Африка и даже Антарктида. В раннем триасе бок о бок с ними добывали себе корм первые лягушки. Позже к ним присоединились первые сухопутные и водные черепахи, а также крокодилы. Очень скоро водные черепахи и крокодилы вторглись в теплые моря. Там они быстро размножились и распространились по всему Мировому океану.
{kind=link}
Первые динозавры были небольшими стройными животными. Поначалу многие из них были больше похожи на птиц, чем на динозавров. Сальтоп ("прыгающая нога") был величиной не больше кошки, а халтикозавр достигал почти 6 м от головы до кончика хвоста. Между этими двумя крайностями существовало множество животных самых разных размеров.
"Собакозубые" и "правящие" рептилии. На суше все еще встречались цинодонты ("сооакозубые") — оыстроногие хищные рептилии, нападавшие на стада медлительных растительноядных животных. Однако появилась и новая группа рептилий — ее называют архозавры, то есть "правящие рептилии". Первые архозавры были маленькими зверьками, которые охотились за мелкой дичью по берегам озер и рек. Впоследствии из них развились куда более крупные животные. В середине триаса возникла еще одна группа рептилий, родственная архозаврам. Это были растительноядные ринхозавры, то есть "клюворылые рептилии". Морда у них оканчивалась довольно странным клювом, которым они, захватывая пищу, пользовались как щипцами. Их челюсти и зубы были отлично приспособлены для того, чтобы резать и рубить твердые растения. Когда пасть ринхозавра закрывалась, нижняя челюсть точно входила в желобок верхней челюсти, как у перочинного ножа, когда его складывают и лезвие вставляется в рукоятку.
{kind=link}
Ящерицеобразная рептилия икарозавр обитала в раннем триасе на территории Северной Америки. У икарозавра были крылья, так сказать, с изменяемой геометрией. Их образовывали участки кожи, туго натянутые на длинные ребра. Когда икарозавр лазил по деревьям, он складывал крылья вдоль тела. А при полете его ребра моментально раздвигались и крылья расправлялись в обе стороны. Они выполняли роль парашюта, на котором животное плавно опускалось на землю.
От текодонтов до динозавров. Ближе к концу триасового периода многие наземные животные, появившиеся в его начале, вымерли. Их место заняли новые рептилии, развившиеся на протяжении триаса. Примерно 225 млн лет назад возникла группа рептилий, которых назвали текодонтами ("ячеистозубыми"). Поначалу это были неуклюжие приземистые животные, немного похожие на крокодилов. Они вели водный образ жизни и плавали при помощи мощного хвоста, загребая при этом задними ногами, которые были намного больше передних. Когда ранние те-кодонты вышли из воды на сушу, их сильные задние ноги быстро приспособились к ходьбе по твердой земле. Вскоре текодонты стали великолепными ходоками и бегунами. Большую часть времени они передвигались по суше на четырех ногах. Однако у них была способность превращаться в истинных спринтеров. Для этого текодонты принимали своего рода "стартовую позу": отклонялись назад, опираясь на свои сверхразвитые задние конечности, и устремлялись вперед на двух ногах, балансируя на бегу длинным хвостом. За последующие 20 млн лет текодонты развились в первых на Земле динозавров.
{kind=link}
На этой схеме представлено внутреннее строение наутилуса. Возможно, схожей внутренней структурой обладали и аммониты.
Еще два важнейших "прорыва". В конце триасового периода в эволюции жизни на Земле произошло еще два важных события. Одно из них случилось на суше и ознаменовалось появлением первых млекопитающих. Второе произошло в воздухе и было связано с пришествием птерозавров ("крылатых рептилий").
{kind=link}
Этот типичный ископаемый аммонит представляет собой плоскую закрученную раковину хорошо различимой спиралевидной формы. Поверхность раковины испещрена узорчатыми ребрами", указывающими на те места, где раньше были перегородки. Глядя на окаменевший аммонит, трудно себе представить, что это останки животного — близкого родственника современных осьминогов и кальмаров. Однако если сравнить его с другим ныне здравствующим головоногим моллюском, наутилусом, то это родство сразу же становится очевидным.
.Пионеры воздухоплавания. Некоторые позвоночные животные уже пытались подняться в воздух. Одним из "первопроходцев" была небольшая ящерица, жившая в пермский период и названная учеными вешельтизавр. Однако у нее не было настоящих крыльев. Она планировала с дерева на дерево на своего рода перепончатом крыле, натянутом между невероятно длинными ребрами. Птерозавры усовершенствовали эту конструкцию и стали первыми позвоночными, которые освоили полноценный полет, будучи сами тяжелее воздуха. У них развилась совершенно иная структура крыла, позволявшая им летать гораздо лучше всех своих предшественников. Удивительные аммониты Аммониты, впервые появившиеся на Земле в девонский период, наиболее изученные и часто встречающиеся ископаемые морские животные. Они принадлежат к группе го-ловоногих моллюсков и, стало быть, являются предшественниками современных осьминогов и кальмаров. Своего наивысшего расцвета, если иметь в виду их численность и разнообразие, аммониты достигли в пермский период. Затем 245 млн лет назад, в конце этого периода, они почти полностью исчезли при массовом вымирании. Но разделаться с аммонитами оказалось не так-то просто. Некоторые из них сумели дожить до триасового периода и благодаря своим склонностям к дальним странствиям по океану, вскоре вновь распространились по всему миру. К середине мезозоя они достигли нового пика эволюционного процветания. Аммониты столь часто встречались в мезозойских морях, а их окаменелости в таком изобилии попадаются в горных породах той эпохи, что они сыграли весьма важную роль в разработке системы идентификации (отождествления) всех морских отложений мезозойской эры. И все же подобное процветание не могло быть вечным, и в конце мелового периода все аммониты внезапно исчезли с лица Земли — вместе со множеством других морских животных, включая белемнитов, плиозавров, ихтиозавров и плезиозавров. От прямых раковин к закрученным У первых головоногих моллюсков, так называемых наутило-идей, были длинные раковины конической формы. Внутри раковин находились газовые камеры, разделенные перегородками. Наутилоидеи выработали весьма примитивный способ втягивания и выталкивания воды, причем выбрасываемую струю они использовали для создания своего рода реактивной тяги. За миллионы лет, прошедшие с тех пор, головоно-гие, включая аммонитов, усовершенствовали этот метод и сделали его своим главным способом передвижения. На протяжении всей палеозойской эры (570—225 млн лет назад) наутилоидеи были наиболее распространенными морскими хищниками. Потом появились новые, оолее высокоорганизованные головоногие, в том числе аммониты, у которых развилась раковина совсем иной формы — завитая, плоская и зачастую с довольно сложным рельефным узором. Перегородки и швы Как и у современных наутилусов, раковина аммонита была разбита на ряд внутренних камер. Каждая из них отделялась от соседней перегородкой. Само животное помещалось в самой последней из новообразованных камер. Места, в которых перегородки присоединялись к раковине, часто хорошо видны на ископаемых останках аммонитов. Их называют шовными линиями. Такие линии образуют на раковинах аммонитов, живших в юрский и меловой периоды, довольно сложные узоры. Ученые используют эти узоры для классификации огромного количества ископаемых аммонитов, обнаруженных за годы исследований. Балласт и плавучесть Раковина аммонита служила как бы балластным (стабилизирующим) механизмом, подобно тому как нынешние наутилусы используют собственные многокамерные раковины. Животное заполняло свободные камеры водой и вновь их опорожняло при помощи особого органа — его называют сифоном. Закачивая и выкачивая воду, аммонит регулировал плавучесть собственного тела, как это делают подводные лодки. Когда животное хотело поглубже погрузиться, оно наполняло свои "балластные отсеки" водой. Когда же оно желало всплыть на поверхность или просто подняться выше, — опорожняло их. Исчезновение аммонитов Невероятное изобилие ископаемых останков аммонитов оказалось несколько обманчивым, заставив ученых в свое время сделать не совсем верные выводы об их распространении в прошлые эпохи. До самого последнего времени специалисты полагали, что аммониты населяли все первобытные моря без исключения. Однако затем было установлено, что отнюдь не все их разновидности обитали по всему земному шару. У разных видов были различные ареалы, что, по всей вероятности, было связано с рядом физических факторов, например с температурой воды или с процентным содержанием в пей соли (соленостью). Смещение материков в конце мелового периода привело к грандиозным климатическим изменениям и общей трансформации (преобразованию) природной среды, затронувшей и Мировой океан. Вероятно, аммониты на сей раз не сумели приноровиться к новым условиям жизни и полностью вымерли.
{kind=link}
Степень закрученности раковин аммонитов различна. Обычно это была форма единой спирали, однако позже у некоторых видов развились частично раскрученные раковины, немного похожие на вопросительный знак с завитушками. Раковины еще более поздних разновидностей по форме смахивали на раковины улиток.
2)Характеристика типа Arthropoda (Членистоногие).
Членистоногие (лат. Arthropoda от греч. ἀρθρον — сустав и греч. πούς/ποδός — нога) — тип первичноротых животных, включающий насекомых, ракообразных, паукообразных и многоножек. По количеству видов и распространённости может считаться самой процветающей группой живых организмов.
Основная особенность представителей данной группы животных организмов - наличие хитинового наружного скелета.
Членистоногие распространены практически всесветно и всюду играют заметную роль. Насекомые, наряду с млекопитающими и птицами, — доминирующая группа наземных животных. Насчитывается не менее 1,5 млн. видов насекомых, возможно значительно больше. Паукообразные и многоножки, хоть и не достигли такого разнообразия, как насекомые, также очень широко распространены. Наконец, ракообразные населяют практически все широты и глубины Мирового океана. В некоторых регионах мелкие ракообразные — господствующая группа планктона.
|
|
Основные особенности строения и биологии
Тело членистоногих сегментировано и разделено на голову, грудь и брюшко. Полностью или частично покрыто хитином. Растут только во время линьки. Конечности членистые; выполняют функции передвижения, дыхания, защиты, захвата и пр. Голова и грудь у некоторых сливается в головогрудь. Пищеварительная система дифференцирована. Ротовой аппарат образован видоизменёнными конечностями. Дыхание жаберное (у ракообразных), лёгочное или трахейное (у паукообразных и насекомых). Кровеносная система незамкнута. Нервная система состоит из головного мозга и брюшной нервной цепочки. Многие членистоногие имеют хорошо развитые органы чувств. Органы выделения представлены метанефридиями и мальпигиевыми сосудами. В основном раздельнополы. Случаи гермафродитизма редки. Развитие протекает с полным или неполным превращением. При неполном метаморфозе у насекомых нет стадии куколки.
Роль членистоногих в природе очень значительна. Они занимают практически все среды обитания и влияют на них. Членистоногие служат пищей животным, опыляют растения, переносят инфекционные заболевания и наносят вред сельскохозяйственным культурам.
Положение группы членистоногих в таксономической иерархии неоднозначно. С одной стороны, некотрыми авторами была предложена теория, согласно которой некоторые подтипы членистоногих не имеют общего предка с кольчатыми червями (Annelida). В частности, предполагалось, что подтип трахейные (Uniramia) ближе к онихофорам (Onychophora), чем к остальным членистоногим. Другими словами, предполагалась полифилия членистоногих. Однако большинство учёных не приняло эту точку зрения, и это также противоречит генетическим исследованиям.
С другой точки зрения, более традиционным вариантом являлось утверждение, что кольчатые черви — ближайшие родственники как членистоногих, так и онихофоров, основываясь на одинаковом количестве сегментов. Позднее некоторые авторы стали говорить о конвергентной эволюции, считая, что членистоногие ближе к нематодам (Nematoda), имея общие с ними способности к линьке. Две вероятные родословные определяют два вышестоящих таксона, к которым можно было бы причислить группу членистоногих животных: членистые (Articulata) и линяющие (Ecdysozoa)*.
Древнейший из известных членистоногих — сприггина (поздний протерозой) — имеет в строении тела немало общего с кольчатыми червями. С другой стороны, не исключается, что сприггина относилась к исчезнувшему типу, хотя и чрезвычайно близкому к членистоногим.
Классификация подтаксонов внутри группы членистоногих также неоднозначна. Пять основных подгрупп причисляют то к подтипам, то к классам. Кроме этих подгрупп имеется также некоторое число ископаемых, в большинстве своём из нижне-кембрийского периода, которые трудно отнести к какой-либо подгруппе либо по причине несхожести с известными группами, либо из-за неясности их родственных взаимоотношений.
Многоножек и насекомых часто объединяют в одну группу неполноусых. Некоторые последние исследования, однако, свидетельствуют, что многоножки не ближе к насекомым, чем к ракообразным.
Здесь мы приводим несколько наиболее известных классификаций, в которых указанные основные группы различным образом объединяются в подтипы (или типы) и надклассы.
Классификация I
Основана на гипотезе о полифилетическом происхождении членистоногих. В связи с этим, тип членистоногих здесь разбивается на три самостоятельных типа.
Тип Pseudognatha Тип Ракообразные Тип Трахейные
Подтип Онихофоры
Подтип Неполноусые
Классификация II
Основана на гипотезе о монофилетическом происхождении членистоногих. В настоящее время общепризнана. При этом тип членистоногих объединяется с онихофорами и тихоходками в группу Lobopoda.
Подтип Трилобитообразные (Trilobitomorpha)(парафилетический таксон)
Подтип Pseudognatha
Надкласс Морские пауки (Pantopoda)
Надкласс Хелицеровые (Chelicerata)
Класс Паукообразные (Arachnida) (пауки, клещи, скорпионы)
Класс Меростомовые (Merostomata) (мечехвосты)
Класс Эвриптериды (Gigantostraca) † (гигантские морские скорпионы)
Подтип Mandibulata
Класс Ракообразные (Crustacea)
Подкласс Remipedia
Подкласс Цефалокариды (Cephalocarida)
Подкласс Жаброногие (Branchiopoda)
Подкласс Ракушковые (Ostracoda)
Подкласс Мистакокариды (Mystacocarida)
Подкласс Веслоногие (Copepoda)
Подкласс Карпоеды (Branchiura)
Подкласс Мешкогрудые (Ascothoracida)
Подкласс Усоногие (Cirripedia)
Подкласс Высшие раки (Malacostraca) (омары, креветки, крабы и др.)
Надкласс Неполноусые (Atelocerata)
Класс Многоножки (Myriapoda)
Подкласс Collifera
Подкласс Скрыточелюстные (Entognatha)
Подкласс Открыточелюстные (Ectognatha) (большая часть насекомых)
ЧЛЕНИСТОНОГИЕ (Arthropoda), крупнейший тип царства животных. Объединяет сегментированных беспозвоночных: пауков, крабов, насекомых и многие другие формы. О том, что особенности организации членистоногих делают их наиболее приспособленными среди всех животных к существованию на нашей планете, говорит то, что один только класс насекомых (Insecta) включает в себя больше видов, чем все прочие группы вместе взятые. Членистоногие адаптированы почти ко всем типам местообитаний и встречаются от высокогорий до океанических глубин. В большинстве групп членистоногих известны паразитические виды; многие членистоногие, особенно насекомые, переносят возбудителей серьезных заболеваний. Некоторые насекомые – злостные вредители, поражающие сельскохозяйственные культуры и леса, портящие пищевые продукты, древесину и другие материалы. Рачки-древоточцы рода Limnoria ежегодно причиняют значительный ущерб, повреждая пирсы, доки и деревянные суда. На борьбу с вредителями и переносчиками болезней, относящимися к типу членистоногих, каждый год тратятся огромные средства, а связанными с ними проблемами занимаются тысячи ученых и полевых работников.
Тело типичного членистоногого состоит из нескольких сегментов, покрытых твердым наружным скелетом (экзоскелетом). У самых примитивных групп все сегменты несут одинаковые членистые придатки. У более эволюционно продвинутых членистоногих придатки на некоторых сегментах отсутствуют и часто превращены в высокоспециализированные структуры, используемые для локомоции, питания, защиты и т.д. У пауков и клещей это четыре пары ходильных ног и придатки, служащие для ловли добычи и питания. Для насекомых характерны три пары ходильных ног и несколько пар придатков с сенсорной и пищедобывающей функцией. Строение ракообразных сильно варьирует, но группа десятиногих раков, к которой относятся, в частности, крабы и омары, отличается пятью парами ходильных ног и многочисленными придатками на голове и брюшке.
Поскольку тело членистоногих покрыто нерастяжимым экзоскелетом, их рост сопровождается периодическими линьками, т.е. сменами покровов. Ставший тесным экзоскелет сбрасывается, и животное увеличивается в размерах, пока не затвердеет новый покров, образовавшийся перед линькой под старым. Экзоскелет секретируется подстилающей его кожей – гиподермой. Многие членистоногие линяют несколько раз. Наступление линек регулируется эндокринной системой, включающей несколько желез или органов.
Кровеносная система членистоногих незамкнутая. Сердце хорошо развито, однако сосуды не образуют сложной сети, и кровь обтекает органы не по тончайшим капиллярам, а выливаясь непосредственно в обширные синусы (пазухи) полости тела. Выделительная система обычно присутствует, но строение ее сильно варьирует и зависит от таксономической группы и среды обитания вида. Если он живет в воде, азотистые и газообразные продукты распада могут диффундировать в окружающую среду непосредственно через поверхность тела. У некоторых членистоногих органы выделения представлены парными нефридиями в некоторых сегментах тела. Головной мозг членистоногих развит сильнее, чем у кольчатых червей, но в остальном нервная система устроена так же, как у этих животных, и состоит из парного вентрального ствола с окологлоточным кольцом и ганглиями, от которых отходят нервы.
Пищеварительный тракт начинается ротовым отверстием и оканчивается анусом. Экзоскелет заходит в них и выстилает переднюю и заднюю части кишки. Эта выстилка при линьке также сбрасывается. Реснички, используемые животными многих других типов для продвижения частиц в кишке или по поверхности тела, у членистоногих полностью отсутствуют.
Способы размножения весьма разнообразны и зависят от среды обитания вида. Яйца членистоногих обычно крупные и богаты желтком. У некоторых сухопутных видов из них вылупляется животное, похожее на миниатюрную взрослую форму, у других – червеобразная личинка. У многих ракообразных личинки дрейфуют в составе планктона и проходят там несколько различных по строению возрастных стадий, прежде чем после очередной линьки превратятся во взрослое животное, способное вести донный образ жизни.
Тип членистоногих делится на три подтипа: трилобитообразные (Trilobita), с единственным классом вымерших трилобитов; челюстные (Mandibulata), с классами Crustacea (ракообразные: морские желуди, омары, крабы, креветки и т.п.), Chilopoda (губоногие), Diplopoda (двупарноногие) и Insecta (насекомые); хелицеровые (Chelicerata) с классами Merostomata (меростомовые: мечехвосты), Pycnogonida (морские пауки) и Arachnida (паукообразные).
Список использованной литературы:
1.Экосистемные перестройки и эволюция биосферы.( А.Г. Понамаренко).
2. Общеобразовательный интернет портал: www.wikipedia.ru
3. Общеобразовательный интернет портал: window.edu.ru
4. Основы Палеонтологии( Т.Г. Кудряшова).
5. Историческая Геология(В. Е.Хаин, Н. В. Короновский, Н.А. Ясаманов).