Контрольная работа: Зрительная система
СОДЕРЖАНИЕ
1 ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЗРИТЕЛЬНОЙ СЕНСОРНОЙ СИСТЕМЫ
1.1 Основные показатели зрения
1.2 Психофизические характеристики света
1.3 Периферический отдел зрительной системы
2 СОМАТОВИСЦЕРАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
2.1 Психофизика кожной механорецепции
2.2 Кожные механорецепторы
2.3 Психофизика терморецепции
2.4 Терморецепторы
2.5 Висцеральная чувствительность
2.6 Проприоцепция
2.7 Функциональный и анатомический обзор центральной соматосенсорной системы
2.8 Передача соматовисцеральной информации в спинном мозгу
2.9 Соматосенсорные функции ствола мозга
2.10 Таламус
2.11 Соматосенсорные проекционные области в коре
2.12 Контроль афферентного входа в соматосенсорной системе
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1 ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ЗРИТЕЛЬНОЙ СЕНСОРНОЙ СИСТЕМЫ
Зрительная система (зрительный анализатор) представляет собой совокупность защитных, оптических, рецепторних и нервных структур, воспринимающих и анализирующих световые раздражители. В физическом смысле свет — это электромагнитное излучение с различными длинами волн — от коротких (красная область спектра) до длинных (синяя область спектра).
Способность видеть объекты связана с отражением света от их поверхности. Цвет зависит от того, какую часть спектра поглощает или отражает предмет. Главные характеристики светового стимула — его частота и интенсивность. Частота (величина, обратная длине волны) определяет окраску света, интенсивность — яркость. Диапазон интенсивностей, воспринимаемых глазом человека — огромен — порядка 1016. Через зрительную систему человек получает более 80% информации о внешнем мире.
1.1 Основные показатели зрения
Зрение характеризуют следующие показатели:
1) диапазон воспринимаемых частот или длин волн света;
2) диапазон интенсивностей световых волн от порога восприятия до болевого порога;
3) пространственная разрешающая способность — острота зрения;
4) временная разрешающая способность — время суммации и критическая частота мельканий;
5) порог чувствительности и адаптация;
6) способность к восприятию цветов;
7) стереоскопия — восприятие глубины.
1.2 Психофизические характеристики света
Психофизические эквиваленты частоты и интенсивности света представлены в таблицах 1.1 и 1.2.
Таблица 1.1. Психофизические эквиваленты частоты света
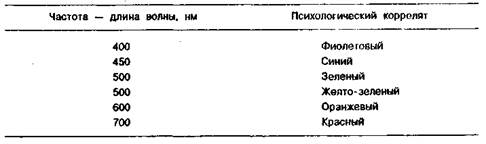
Таблица 1.2. Психофизические эквиваленты интенсивности света
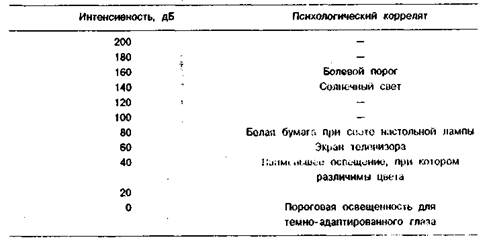
Для характеристики восприятия света важны три качества: тон, насыщенность и яркость. Тон соответствует цвету и меняется с изменением длины волны света. Насыщенность означает количество монохроматического света, добавление которого к белому свету обеспечивает получение ощущения, соответствующего длине волны добавленного монохроматического света, содержащего только одну частоту (или длину волны). Яркость света связана с его интенсивностью. Диапазон интенсивностей света от порога восприятия до величин, вызывающих болевые ощущения, огромен — 160 дБ. Воспринимаемая человеком яркость объекта зависит не только от интенсивности, но и от окружающего его фона. Если фигура (зрительный стимул) и фон освещены одинаково, то есть между ними нет контраста, яркость фигур возрастает с увеличением физической интенсивности освещения. Если контраст между фигурой и фоном увеличивается, яркость воспринимаемой фигуры уменьшается с увеличением освещенности.
Пространственная разрешающая способность — острота зрения — минимальное различимое глазом угловое расстояние между двумя объектами (точками). Острота определяется с помощью специальных таблиц из букв и колец и измеряется величиной I/a, где а — угол, соответствующий минимальному расстоянию между двумя соседними точками разрыва в кольце. Острота зрения зависит от общей освещенности окружающих предметов. При дневном свете она максимальна, в сумерках и в темноте острота зрения падает.
Временные характеристики зрения описываются двумя основными показателями — временем суммации и критической частотой мельканий.
Зрительная система обладает определенной инерционностью: после включения стимула необходимо время для появления зрительной реакции (оно включает время, требующееся для развития химических процессов в рецепторах). Исчезает зрительное впечатление не сразу, а лишь через некоторое время после прекращения действия на глаз света или изображения, поскольку для восстановления зрительного пигмента сетчатке глаза также требуется время. Существует эквивалентность между интенсивностью и длительностью действия света на глаз. Чем короче зрительный стимул, тем большую интенсивность он должен иметь, чтобы вызывать зрительное ощущение. Таким образом, для возникновения зрительного ощущения имеет значение суммарное количество световой энергии. Эта связь между длительностью и интенсивностью сохраняется лишь при коротких длительностях стимулов — до 20 мс. Для более длительных сигналов (от 20 мс до 250 мс) полная компенсация пороговой интенсивности (яркости) за счет длительности уже не наблюдается. Всякая зависимость между способностью к обнаружению света и его длительностью исчезает после того, как продолжительность стимула достигает 250 мс, а при больших длительностях решающей становится интенсивность. Зависимость пороговой интенсивности света от длительности его воздействия называется временной суммацией. Этот показатель используется для оценки функции зрительной системы.
Зрительная система сохраняет следы светового раздражения в течение 150-250 мс после его включения. Это свидетельствует о том, что глаз воспринимает прерывистый свет, как непрерывный, при определенных интервалах между вспышками. Частота вспышек, при которой ряд последовательных вспышек воспринимается как непрерывный свет, называется критической частотой мельканий. Этот показатель неразрывно связан с временной суммацией: процесс суммации обеспечивает плавное слияние последовательных изображений в непрерывный поток зрительных впечатлений. Чем выше интенсивность световых вспышек, тем выше критическая частота мельканий. Критическая частота мельканий пи средней интенсивности света составляет 16-20 в 1 с.
Порог световой чувствительности — это наименьшая интенсивность света, которую человек способен увидеть. Она составляет 10 -10 — 10 -11 эрг/с. В реальных условиях на величину порога существенно влияет процесс адаптации — изменения чувствительности зрительной системы в зависимости от исходной освещенности. При низкой интенсивности света в окружающей среде развивается темповая адаптация зрительной системы. По мере развития темновой адаптации чувствительность зрения возрастает. Длительность полной темновой адаптации составляет 30 мин. При увеличении освещенности окружающей среды происходит световая адаптация, которая завершается за 15-60 с. Различия темновой и световой адаптации связаны со скоростью химических процессов распада и синтеза пигментов сетчатки.
Восприятие света зависит от длины волны света, попадающего в глаз. Однако, такое утверждение справедливо лишь для монохроматических лучей, то есть лучей с одной длиной волны. Белый свет содержит все длины световых волн. Существует три основных цвета: красный — 700 нм, зеленый — 546 нм и синий — 435 нм. В результате смешивания основных цветов можно получить любой цвет. Объясняют цветовое зрение на основе предположения о существовании в сетчатке глаза фоторецепторов трех различных типов, чувствительных к различным длинам волн света, соответствующих основным частотам спектра (синий, зеленый, красный).
Нарушение восприятия цвета называется цветовой слепотой, или дальтонизмом, по имени Дальтона, который впервые описал этот дефект зрения на основе собственного опыта. Дальтонизмом страдают, в основном, мужчины (около 10%) в связи с отсутствием определенного гена в Х-хромосоме. Известны три типа нарушений светового зрения: протанопия — отсутствие чувствительности к красному цвету, дейтеранопия — отсутствие чувствительности к зеленому цвету и тританопия — отсутствие чувствительности к синему цвету. Полная цветовая слепота — монохроматия — встречается исключительно редко.
Бинокулярное зрение — участие обоих глаз в формировании зрительного образа — создается за счет объединения двух монокулярных изображений объектов, усиливая впечатление пространственной глубины. Поскольку глаза расположены в разных "точках" головы справа и слева, то в изображениях, фиксируемых разными глазами, имеются небольшие геометрические различия (диспарантность), которые тем больше, чем ближе находится рассматриваемый объект. Диспарантность двух изображений лежит в основе стереоскопии, то есть восприятия глубины. Когда голова человека находится в нормальном положении, возникают отклонения от точно соответствующих проекций изображений в правом и левом глазах, так называемая диспарантность рецептивных полей. Она уменьшается с увеличением расстояния между глазами и объектом. Поэтому на больших расстояниях между стимулом и глазом глубина изображения не воспринимается.
1.3 Периферический отдел зрительной системы
Снаружи глаз виден как сферическое образование, прикрытое верхним и нижним веком и состоящее из склеры, коньюктивы, роговицы, радужной оболочки. Склера представляет собой соединительную ткань белого цвета, окружающую глазное яблоко. Коньюктива — прозрачная ткань, снабженная кровеносными сосудами, которая на переднем полюсе глаза соединяется с роговицей. Роговица является прозрачным защитным наружным образованием, кривизна поверхности которого определяет особенности преломления света. Так, при неправильной кривизне роговицы возникает искажение зрительных изображений, называемое астигматизмом. Позади роговицы находится радужная оболочка, цвет которой зависит от пигментации составляющих ее клеток и их распределения. Между роговицей и радужной оболочкой находится передняя камера глаза, наполненная жидкостью — "водянистой влагой". В центре радужной оболочки находится зрачок круглой формы, пропускающий внутрь глаза свет после его прохождения через роговицу.
Размер зрачка зависит от освещенности. Контроль за изменениями размера зрачка осуществляется автоматически нервными волокнами, заканчивающимися в мускулатуре радужной оболочки. Круговая мышца, суживающая зрачок — сфинктер — иннервируется парасимпатическим волокнами, мышца, расширяющая зрачок — дилататор — иннервируется симпатическими волокнами. Изменения диаметра зрачка меняют интенсивность светового раздражения незначительно — всего в 16-17 раз (если учитывать, что диапазон интенсивности света изменяется в 16 млрд. раз). Реакция расширения зрачка до максимального диаметра — 7,5 мм — очень медленная: она длится около 5 минут. Максимальное сокращение диаметра зрачка до 1,8 мм достигается быстрее — всего за 5 секунд. Это значит, что основная функция зрачка состоит не в регуляции интенсивности света вообще, а в том, чтобы пропускать лишь тот свет, который попадает на центральную часть хрусталика, где фокусировка наиболее точная. Сужение зрачка направлено на сохранение наиболее возможной при данных условиях освещенности глубины резкости.
Роговица и коньюктива покрыты тонкой пленкой слезной жидкости, секретируемой в слезных железах, расположенных в височной части глазницы, над глазным яблоком. Слезы защищают роговицу и коньктиву от высыхания.
Позади радужной оболочки расположены задняя камера глаза и хрусталик. Хрусталик — двояковыпуклая линза, расположенная в сумке, волокна которой соединены с ресничными мышцами и наружным сосудистым слоем сетчатки. Хрусталик может становиться более плоским или более выпуклым в зависимости от расстояния между глазом и объектом. Изменение кривизны хрусталика называется аккомодацией. Внутри глаза, позади хрусталика, находится стекловидное тело. Оно представляет собой коллоидный раствор гиалуроновой кислоты во внеклеточной жидкости.
Аккомодация хрусталика иногда оказывается недостаточной, чтобы спроецировать изображение точно на сетчатку. Если расстояние между хрусталиком и сетчаткой больше, чем фокусное расстояние хрусталика, то возникает близорукость (миопия). Если сетчатка расположена слишком близко к хрусталику и фокусировка хороша только при рассматривании далеко расположенных предметов, возникает дальнозоркость (гиперметропия). Близорукость и дальнозоркость корректируются очками с вогнутыми и выпуклыми линзами соответственно.
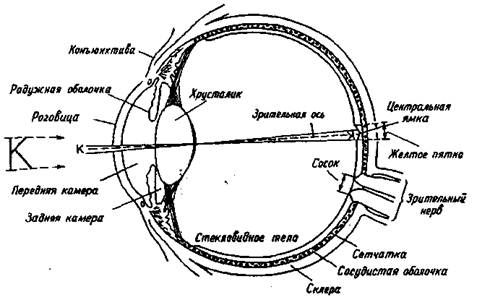
Рис. 1.1. Горизонтальный срез правого глаза
Астигматизм (результат неравномерной кривизны роговицы) плохо корректируется даже сложными линзами. Для его исправления более пригодны контактные линзы, которые, плавая в слезной жидкости над роговицей, компенсируют ее отклонения от правильной формы. Итак, оптическая система глаз обеспечивает фокусировку изображения на рецепторной поверхности сетчатки. Диоптрический аппарат, состоящий из системы линз, передает на сетчатку резко уменьшенное изображение предметов (рис. 1.1).
Сетчатка — с нейроанатомической точки зрения — высокоорганизованная слоистая структура, объединяющая рецепторы и нейроны (рис. 1.2). Фоторецепторные клетки — палочки и колбочки — расположены в пигментном слое, наиболее удаленном от хрусталика.
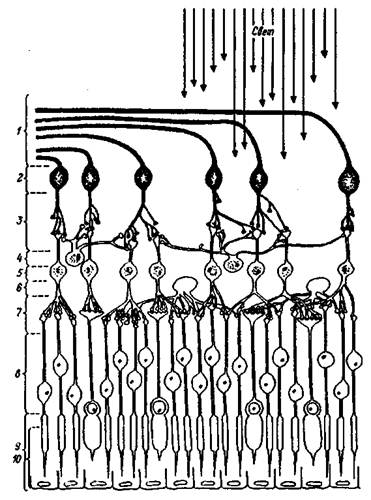
Рис. 1.2. Строение сетчатки глаза
Вверху — падающий свет; 1 — волокна зрительного нерва; 2 — ганглиозные клетки; 3 — внутренний синаптический слой; 4 — амакриновые клетки; 5 — биполярные клетки; 6 — горизонтальные клетки; 7 — наружный синаптический слой, 8 — ядра рецепторов; 9 — рецепторы; 10 — пигментный слой эпителиальных клеток.
Они повернуты от пучка падающего света таким образом, что их светочувствительные концы спрятаны в промежутках между сильно пигментированными эпителиальными клетками. Эпителиальные пигментные клетки участвуют в метаболизме фоторецепторов и синтезе зрительных пигментов. Все нервные волокна, выходящие из сетчатки, лежат в виде переплетенного пучка на пути света, создавая препятствие на пути его попадания в рецепторы. Кроме того, в том месте, где они выходят их сетчатки по направлению к мозгу, отсутствуют светочувствительные элементы — это так называемое слепое пятно. Свет, попадающий на сетчатку в области слепого пятна не воспринимается элементами сетчатки, поэтому остается "дефект" изображения, проецируемого на сетчатку. Однако, наличие слепого пятна не сказывается на целостности зрительного восприятия. Этот эффект или, точнее, дефект слепого пятна компенсируют высшие зрительные центры.
Палочки и колбочки отличаются как структурно, так и функционально. Зрительный пигмент (пурпур — родопсин) — содержится только в палочках. В колбочках находятся другие зрительные пигменты — иодопсин, хлоролаб, эритлаб, необходимые для цветового зрения. Палочка в 500 раз более чувствительна к свету, чем колбочка, но не реагирует на свет с разной длиной волны, т.е. она не цветочувствительна. Зрительные пигменты расположены в наружном сегменты палочек и колбочек. Во внутреннем сегменте находится ядро и митохондрии, принимающие участие в энергетических процессах при действии света.
В глазу человека около 6 млн. колбочек и 120 млн. палочек — всего около 130 млн. фоторецепторов. Плотность колбочек наиболее высока в центре сетчатки и падает к периферии. В центре сетчатки, в небольшом ее участке, находятся только колбочки. Этот участок называется центральной ямкой. Здесь плотность колбочек равна 150 тысячам на 1 квадратный миллиметр, поэтому в области центральной ямки острота зрения максимальна. Палочек в центре сетчатки очень мало, их больше на периферии сетчатки, но острота "периферического" зрения при хорошей освещенности невелика. В условиях сумеречного освещения преобладает периферическое зрение, а острота зрения в области центральной ямки падает. Таким образом, колбочки функционируют при ярком свете и выполняют функцию восприятия цвета, палочкой воспринимают свет и обеспечивают зрительное восприятие при слабой освещенности.
Первичный процесс зрительной рецепции — фотохимическая реакция. Фотоны поглощаются молекулами зрительных пигментов. Каждая молекула пигмента поглощает один фотон (квант света) и переходит на более высокий энергетический уровень. Поглощение кванта света в фоторецепторе запускает многоступенчатый процесс распада молекул пигмента. Родопсин — зрительный пигмент палочек — состоит из белка (опсина) и ретиналя (альдегида витамина А1). При распаде родопсина образуются опсин и витамин А1. Иодопсин — основной пигмент колбочек — также состоит из опсина и ретиналя. Фотохимические процессы в палочках и колбочках сходны. Родопсин и иодопсин имеют разные спектры поглощения: максимум спектра поглощения родопсина — 500 им (зелено-голубая часть), максимум спектра иодопсина — 570 нм (желтая часть). Каждая палочка в сетчатке человека содержит один пигмент, каждая колбочка — три разных пигмента, максимумы поглощения которых составляют примерно 425, 435 и 570 нм. Восстановление пигментов осуществляется в темноте в результате цепи химических реакций (ресинтез), протекающих с поглощением энергии. Ретиналь ресинтезируется на основе цис-изомера витамина А1, поэтому при недостатке витамина А1 в организме возникает недостаточность сумеречного зрения. Если освещение постоянно и равномерно, то фотохимический распад пигментов находится в равновесии с их ресинтезом. Этот фотохимический процесс обеспечивает светотемновую адаптацию.
При освещении фоторецептора возникает увеличение элетроотрицателъности потенциала внутри клетки по отношению к внеклеточному пространству. Это приводит к уменьшению трансмембранного тока в рецепторах. Таким образом, на свет в фоторецепторах возникает гиперполяризационный ответ. Гиперполяризация отличает зрительные рецепторы от других рецепторов, например, слуховых и вестибулярных, где возбуждение связано с деполяризацией мембраны. Амплитуда рецепторного зрительного потенциала увеличивается при увеличении интенсивности света (освещенности, относительно предыдущего состояния адаптации). Амплитуда рецепторного потенциала зависит также от длины волны света, максимум ответа палочек проявляется при длине волны максимального поглощения родопсина — 500 нм, колбочек — 560-570 нм.
Палочки и колбочки соединены с биполярными нейронами сетчатки, которые, в свою очередь, образуют с ганглиозными клетками синапсы, выделяющие ацетилхолин. Аксоны ганглиозных клеток сетчатки в составе зрительного нерва идут к различным мозговым структурам. Около 130 млн. фоторецепторов связаны с 1,3 млн., волокон зрительного нерва, что свидетельствует о конвергенции зрительных структур и сигналов. Только в центральной ямке каждая колбочка связана с одной биполярной клеткой, а она, в свою очередь, — с одной ганглиозной. К периферии от центральной ямки на одной биполярной клетке конвергируют множество палочек и несколько колбочек, а на ганглиозной — множество биполярных. Поэтому функционально такая система обеспечивает переработку первичного сигнала, повышающую вероятность его обнаружения за счет широкой конвергенции связей от периферических рецепторов к ганглиозной клетке, посылающей сигналы в мозг (рис. 1.3).
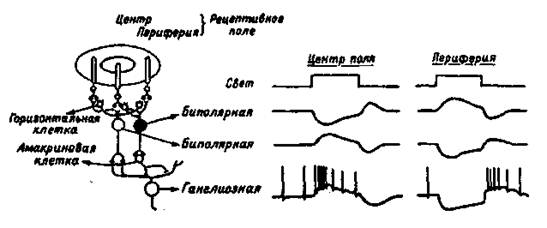
Рис. 1.3. Организация концентрических рецептивных полей биполярных и ганглиозных клеток сетчатки (слева) и схема изменений потенциала, построенная на основе внутриклеточных записей (справа).
Два тина тормозных нейронов — горизонтальные и амакриновые клетки — расположены в том же слое, где находятся биполярные нейроны, ограничивают распространение зрительного возбуждения внутри сетчатки. Горизонтальные и амакриновые клетки связаны с биполярными и ганглиозными горизонтальными связями, обеспечивающими латеральное торможение между соседними клеточными элементами сетчатки: горизонтальные — между биполярными, амакриновые — между ганглиозными.
Ганглиозные клетки сетчатки при слабой освещенности дают непрерывную импульсацию. При усилении освещенности половина клеток усиливает импульсацию, половина — ослабляет. Следовательно, первые являются детекторами яркости, вторые — детекторами темноты. Все ганглиозные клетки сетчатки имеют круглые рецептивные поля в отличие от неправильных нессиметричных рецептивных полей слуховых и соматических нейронов. Оптимальным стимулом для ганглиозных клеток служит либо светлое пятно, окруженное темным поясом, либо темное пятно, окруженное ярким поясом. Многие ганглиозные клетки реагируют только на изменение освещения, но не реагируют на постоянный свет. Схематическое изображение рецептивных полей и реакций нейронов сетчатки показано на рис.1.3.
Каждая ганглиозная клетка имеет свое рецептивное поле, т.е. ограниченный участок сетчатки, фоторецепторы которого связаны с данной клеткой. Реакция такой клетки на свет вне ее рецептивного поля отсутствует. Ганглиозные клетки функционально различны, их разделяют на два типа:
1) нейроны, которые возбуждаются светом, падающим на центр рецептивного поля, но затормаживаются, если свет падает на его периферию;
2) нейроны, которые затормаживаются светом в центре рецептивного поля и возбуждаются при действии света на его края.
Одновременная реакция нейронов первого и второго типов лежит в основе появления одновременного контраста за счет подчеркивания края изображения их антагонистическими рецептивными полями.
Размеры рецептивных полей, равно как и их функциональные свойства, зависят от расположения фоторецепторов данного поля на сетчатке относительно центральной ямки (эксцентриситет). Величина рецептивных полей растет от области центральной ямки к периферии сетчатки. Это является следствием структурной организации связей элементов сетчатки. Так, в середине сетчатки, в области центральной ямки, где плотность колбочек максимальная, одна колбочка через отдельную биполярную клетку соединяется с отдельной ганглиозной клеткой. Таким образом, рецептивные поля ганглиозных клеток, связанные с центром сетчатки очень узкие и не перекрываются. На периферии, где, в основном, находятся палочки, отмечаются широкие рецептивные поля: множество рецепторов связано с одной ганглиозной клеткой.
Функционально острота зрения зависит и от свойств рецептивных полей: острота зрения — возможность различения двух соседних точек — максимальна для узких рецептивных полей центральной ямки. В то же время слабые сигналы с периферии сетчатки выделяются зрением, благодаря взаимодействию широких перекрывающихся рецептивных полей за счет пространственной суммации раздражителей.
При освещении сетчатка генерирует электрические потенциалы, которые называют электроретинограммой. Эта суммарная электрическая реакция отражает процессы возбуждения различных нервных структур: волна возникает во внутренних сегментах фоторецепторов; волна является результатом возбуждения биполярных и амакриновых клеток сетчатки; волна связана с пигментным эпителием; волна, является реакций горизонтальных клеток сетчатки на выключение света.
Таким образом, волны электроретинограммы имеют своим источником все клеточные элементы сетчатки, кроме ганглиозных.
В сетчатке обнаружены клетки, для которых наиболее эффективны цвета-антагонисты красный и зеленый, а также клетки, для которых пару антагонистов составляют желтый и синий или зеленый и синий. Объяснение антагонистического действия цветов на ганглиозные клетки состоит в том, что из трех типов колбочек два всегда связаны с одним нейроном, а часть колбочек имеет возбудительные синапсы, другая часть — тормозные.
Сетчатку, по сложности организации, часто рассматривают как часть мозга, расположенную на периферии. Здесь с фоторецепторами связаны несколько слоев нейронов, формирующих афферентный поток, который идет к подкорковым и корковым центрам зрительной системы. Горизонтальные и биполярные клетки сетчатки не генерируют потенциалов действия, основной формой их активности являются градуальные гиперполяризации и деполяризации. Ганглиозные клетки генерируют потенциалы действия, которые проводятся по их длинным аксонам, составляющим зрительный нерв.
Зрительный нерв содержит около 800 тысяч волокон ганглиозных клеток сетчатки. Зрительные нервы обоих глаз перекрещиваются в области основания черепа, где около полумиллиона волокон зрительного нерва переходят на противоположную сторону. Остальные 300 тысяч волокон вместе с перекрещенными аксонами второго зрительного нерва образуют зрительный тракт.
Нервные волокна зрительного тракта подходят к четырем структурам мозга: ядрам верхних бугров четверохолмия — средний мозг; ядрам латерального коленчатого тела — таламус; супрахиазмальным ядрам гипоталамуса; к глазодвигательным нервам.
Ядра верхних бугров четверохолмия и латерального коленчатого тела являются конечными пунктами двух параллельных путей от ганглиозных клеток сетчатки: одна ветвь аксона ганглиозной клетки идет в латеральное коленчатое тело, другая — в верхнее двухолмие. Обе ветви сохраняют упорядоченную проекцию сетчатки. От переднего двухолмия после переключения сигналы идут к крупному ядру таламуса — подушке.
Аксоны клеток латерального коленчатого тела, проходящие в составе зрительной радиации, проецируются к клеткам первичной зрительной коры (поле 17 или стриарная кора). Проекция зрительной ямки сетчатки — зоны максимальной остроты зрения — в 35 раз больше проекции участка такого же размера на периферии сетчатки. Клетки поля 17 (стриарной коры) связаны с полями 18 и 19 (престриарная кора), так называемыми вторичными зрительными зонами. От этих зон идут проекции к подушке таламуса, куда поступает информация от верхних бугров четверохолмия. Кроме того, зрительные пути прослеживаются к лобной коре, они примыкают к ассоциативной коре.
Клетки латерального коленчатого тела, получающие основную афферентацию от сетчатки, имеют простые концентрические рецептивные поля, как и ганглиозные клетки. Здесь проявляется бинокулярное взаимодействие: волокна от обоих глаз распределены топографически правильно, послойно. В то же время небольшая часть клеток латерального коленчатого тела активируется от обоих зрительных нервов.
Нейроны зрительной коры уже имеют не концентрические, а почти прямоугольные зрительные поля, некоторые из нейронов реагируют на определенную ориентацию (наклон) полосы — светлой или темной (рис. 1.4).
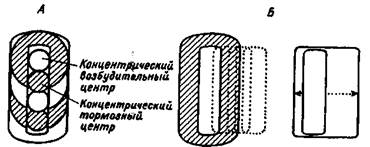
Рис. 1.4. Концентрические рецептивные поля в сетчатке и подкорковых зрительных центрах (А), прямоугольные и сложные рецептивные поля в зрительной коре (Б).
В зрительной коре существуют два функционально различных типа клеток: простые и сложные. Простые клетки имеют рецептивное поле, состоящее из возбудительной и тормозной зоны, которые можно предсказать на основе исследования реакции клетки на маленькое световое пятно. Структуру рецептивного поля сложной клетки невозможно установить сканированием светового пятнышка. Они служат "детекторами" угла, наклона или движения линий в поле зрения. В коре уже совершенно отчетлива бинокулярная конвергенция: в одной точке представлены симметричные поля зрения — справа и слева.
Близко расположенные клетки зрительной коры "видят" только небольшую часть поля зрения. Лежащие друг под другом нейроны одной колонки коры реагируют на один и тот же стимул, оптимальный по ориентации, наклону и направлению движения. В одной колонке могут располагаться как простые, так и сложные клетки. Простые клетки найдены в III и IV слоях, где заканчиваются таламические волокна. Сложные клетки расположены в более поверхностных слоях коры 17 поля. В полях 18 и 19 зрительной коры простые клетки являются исключением, здесь расположены сложные и сверхсложные клетки. Последние реагируют, например, только на стимулы определенной ширины, длины и ориентации.
Итак, от уровня к уровню зрительной системы происходит усложнение рецептивных полей нейронов. Все рецептивные поля организованы в виде возбудительных и тормозных зон. Концентрические рецептивные поля, характерные для сетчатки и латерального коленчатого тела, уже не встречаются в коре. В зрительной системе, как и в других сенсорных системах, чем выше синаптический уровень, тем строже ограничены функции отдельных нейронов — детекторов свойств.
Для успешной работы системы распознавания зрительных образов очень важны движения глаз. Известно, что глаз человека приводится в движение шестью наружными мышцами. Относительно координат головы глаза двигаются горизонтально, вертикально и вокруг своей оси. Если оба глаза двигаются в одном направлении, такие движения называются содружественными. При переводе взгляда с ближней точки на дальнюю осуществляются дивергентные движения. При наклоне головы в сторону наблюдаются небольшие вращательные движения глаз.
При взгляде на любой предмет глаза двигаются от одной точки фиксации к другой быстрыми скачками — саккадами. Длительность саккад от 10 до 80 мс, длительность периодов фиксации 150-300 мс. Медленные движения глаз реализуются при слежении за движущимися объектами — следящие движения.
Движения глаз управляются центрами, которые находятся в области ретикулярной формации мозга и среднего мозга, в верхних буграх четверохолмия и в претектальной области. Все эти подкорковые центры координируются сигналами из зрительной, теменной и лобной коры, ответственными за программирование движений тела и оценки его положения в пространстве. Для наиболее тонкой регуляции глазодвигательных функций весьма существенны влияний мозжечка, сравнивающего тонический и фазный компоненты движения при ориентации и пространстве.
В процессе зрительного восприятия, особенно при слежении за движущимся объектом, возникает оптический нистагм, вызываемый движущимися оптическими стимулами и состоящий из чередовании саккад и медленные следящих движений. Движения глаз имеют огромное значение для восприятия: при неподвижном глазном яблоке восприятие изображения пропадает в связи с разложением пигмента и адаптацией фоторецепторов.
Координированные движения глаз обеспечивают объединение информации, идущей от обоих глаз в центры мозга. Особое значение для восприятия и координации движений играют нейроны переднего двухолмия. Они организованы в колонки, которые воспринимают сигналы, поступающие от одних и тех же участков полей зрения: активность нейронов мою отдела мозга, на которых конвергирует импульсация от правого и левого глаза, является пусковым механизмом для глазодвигательных нейронов. На высших уровнях зрительной системы параллельно функционируют две системы анализа: одна определяет место предмета и пространстве, другая описывает его признаки. Конечные результаты параллельных процессов интегрируются и возникает законченный зрительный образ внешнего предметного мира.
2 СОМАТОВИСЦЕРАЛЬНЫЕ ВЗАИМОДЕЙСТВИЯ
2.1 Психофизика кожной механорецепции
Пороги субъективная интенсивность тактильных стимулов
Абсолютный порог для тактильного стимула можно измерить калиброванными волосками по методу, который ввел в психофизику в XIX в. немецкий физиолог Макс фон Фрей (Max von Frey). В современных стимуляторах высокоточные движения раздражающего стерженька обеспечиваются электромагнитом. Стерженек обеспечивает воздействие градуированной интенсивности с заданной временной характеристикой, например одиночные импульсы или ритмичные нажимы с синусоидально изменяющейся интенсивностью (вибрации).
Пороги восприятия, определенные волосками Фрея и вибрациями (200 Гц), генерируемыми электромагнитным стимулятором, приведены для разных частей тела в табл. 2.1.
Таблица 2.1. Пороги восприятия механической стимуляции кожи человека
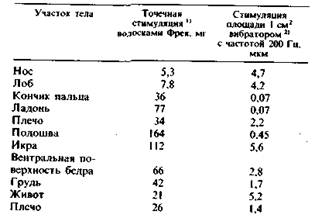
Кончики пальцев воспринимают вибрацию амплитудой менее 1 мкм. Относительная чувствительность разных частей тела весьма неодинакова. Кроме того, разные по своей природе стимулы демонстрируют неодинаковую чувствительность одного и того же места (табл. 2.1). Это объясняется тем, что околопороговая вибрация и волоски Фрея стимулируют разные типы механорецепторов.
Пространственное разрешение тактильных стимулов
Для получения меры пространственного paзрешения пригодны различные тесты по распознаванию пространственных деталей тактильных стимулов. В одном из часто применяемых тестов используя циркуль, двумя кончиками которого одновременно прикасаются к коже (рис. 2.1) для определения двухточечного порога.
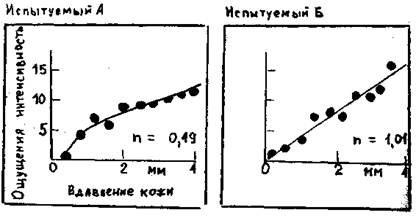
Рис. 2.1. Психофизические опыты для оценки интенсивности стимула. Испытуемый выражает ощущаемую им силу давления на ладонь (здесь – длительностью 1с.) числом на произвольно выбранной шкале (ось ординат). Объективную интенсивность стимула (ось обсцисс) меняли в случайном порядке. Кривые – степенные функции, соответствующие полученным данным с показателем степени n, отмеченным графиках.
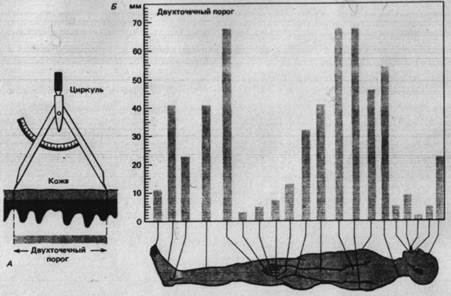
Рис. 2.2. Двухточечный порог кожи. А. Кончики циркуля, разведенные на разные расстояния, неоднократно прикасаются к коже. Двухточечный порог - наименьшее расстояние, при котором испытуемый может определить, чтo стимулируются две точки, а не одна. Б. Изменчивость двухточечного порога кожи в зависимости oт участка тела человека.
Испытуемый должен, не глядя, ответить, в одной или в двух точках до него дотронулись. Двухточечный порог на разных частях тела неодинаков (рис. 2.2 Б). Это обусловлено особенности иннервации разных областей кожи и связанным с ней характером соединения афферентных волокон в ЦНС.
2.2 Кожные механорецепторы
Исследования на человеке и животных привели к хорошему пониманию физиологической функции гистологического строения механорецепторов кожи и обнаружили между ними корреляцию. Кожа млекопитающих содержит несколько основных типов механорецепторов.
Типы низкопороговых механорецепторов кожи
Нейрофизиологические
опыты показали, что лишенная волос кожа крыс, кошек, обезьян и людей одержит
три основных типа механорецепторов с афферентами группы II (А![]() ). Их называют рецепторами
МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном
воздействии механического стимула (давления; например, давления веса тела на
подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток
импульсов. БА – это быстро адаптирующийся рецептор, реагирующий только на
механические стимулы, которые изменяются во времени. ТП означает тельца Пачини,
гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся
механорецепторы.
). Их называют рецепторами
МА, БА и ТП. МА означает медленно адаптирующийся: при долговременном
воздействии механического стимула (давления; например, давления веса тела на
подошву) такой рецептор посылает по своему афферентному волокну непрерывный поток
импульсов. БА – это быстро адаптирующийся рецептор, реагирующий только на
механические стимулы, которые изменяются во времени. ТП означает тельца Пачини,
гистологически выявленные структуры, функционирующие как очень быстро адаптирующиеся
механорецепторы.
Рецептивные поля и плотность иннервации механорецепторов
Область, в которой стимул данной интенсивности способен возбудить механочувствительное афферентное волокно, называется его рецептивным полем. Оно приблизительно соответствует анатомической протяженности всех окончаний этого волокна. Например, один афферент связан с двумя-тремя тактильными тельцами оволосненной кожи, а все 30-50 дисков Меркеля такого тельца иннервируются одной коллатералью. Концевые разветвления гораздо протяженнее у рецепторов волосяных фолликулов: каждый афферент может отходить от многих фолликулов, и каждый фолликул иннервируется несколькими афферентными волокнами.
Какие свойства рецепторов определяют пространственное разрешение, измеряемое двухточечным порогом? Согласно результатам различных исследований, величина рецептивного поля здесь не главное; решающую роль играет плотность иннервации (т. е. число афферентных волокон на 1 см2 поверхности кожи). Валлбо с сотрудниками, проведя микронейрографические измерения у людей, пришли к выводу, что пространственному разрешению на разных участках кисти соответствует только плотность афферентов БА и MA. Поскольку человек, как известно, лучше всего распознает предметы на ощупь подвижными пальцами, можно предположить главную роль в этом рецепторов БА.
2.3 Психофизика терморецепции
Терморецепция (температурная чувствительность) кожи включает два качественных типа – чувство холода и чувство тепла. Известно, что в коже человека есть специфические холодовые и тепловые точки, в каждой из которых можно вызвать ощущение только холода или только тепла. Холодовых точек больше, чем тепловых.
Например, на 1 см2 ладони 1-5 холодовых и только 0,4 тепловых. Наибольшая плотность этих точек обнаружена в самой чувствительной к температуре области – на лице; здесь на 1 см2 кожи 16-19 холодовых точек, а одиночных сенсорных точек, реагирующих на тепло, не обнаруживается. Восприятие температурных стимулов, включая сопровождающие их регуляторные реакции, обладает аффективным действием. Иными словами, они могут казаться приятными или неприятными: человек может мерзнуть или чувствовать освежающую прохладу, согреваться или мучиться от жары.
Психофизическое исследование терморецепции
Существует температурный диапазон, в котором при постоянстве температурного стимула мы не ощущаем ни тепла, ни холода; иначе говоря, в этой игральной зоне температурная чувствительность полностью адаптирована. За пределами нейтральной зоны устойчивые температурные ощущения возникают даже при постоянной температуре (ноги могут мерзнуть часами). Верхний и нижний пределы нейтральной зоны для участка кожи площадью 15 см2 равны соответственно 36 и 30 °С.
Устойчивые ощущения тепла и холода
Продолжительное ощущение тепла при температуре кожи выше 36 °С тем сильнее, чем выше эта температура. При температуре около 45 °С чувство тепла сменяется болью от горячего. Когда обширные области охлаждаются до температуры ниже 30 °С, возникает устойчивое ощущение холода; боль от холода возникает при температуре кожи 17 °С и ниже.
Динамические температурные ощущения
Температурные ощущения, испытываемые при изменении температуры кожи, в основном определяются тремя параметрами: ее исходной температурой, скоростью изменения последней и размерами участка, на который действует стимул.
Прохладную кожу (скажем, при 28 °С) нужно только слегка охладить (меньше чем на 0,2 °С), чтобы превратить устойчивое чувство холода в ощущение «стало холоднее». Но тот же участок кожи надо нагреть почти на 1 °С для появления чувства тепла. Аналогичным образом, при исходной температуре кожи 38 °С уже незначительное нагревание (менее чем на 0,2 С) вызывает ощущение «стало теплее», между тем как для появления чувства холода надо охладить кожу приблизительно на 0,8 °С.
Если скорость изменения температуры превышает примерно 5 °С/мин, это мало влияет на тепловой или холодовой пороги; при более медленных ее изменениях оба будут постепенно повышаться. Например, если кожа охлаждается на 0,4°С/мин, начиная от температуры 33,5 °С, для появления чувства холода понадобится 11 мин; за это время температура упадет на 4,4 °С. Когда охлаждение идет очень медленно, человек может не заметить, как обширные участки кожи стали совсем холодными (при одновременной потере тепла телом), особенно если его внимание отвлечено чем-то другим. Предположительно этот фактор действует, когда человек простужается.
2.4 Терморецепторы
Реакции при постоянной температуре кожи
При переходе примерно от 30 до 43 °С частота импульсации у тепловых рецепторов возрастает, а затем резко падает; в пороговой области ощущения боли от горячего (приблизительно 45 °С) тепловые рецепторы обычно невозбудимы. Частота импульсации холодовых рецепторов повышается по мере падения температуры от 33-40°С и достигает максимума при 17-26 °С (у разных рецепторов). Из-за этого максимума каждая частота разрядов связана с двумя температурами, но тем не менее ЦНС может правильно распознавать холод и на отрезке ниже 17°С.
Реакции при изменении температуры кожи
Сравнение показывает, что частота импульсации зависит не только от температуры (пропорциональный рецептор), но и от скорости ее изменения (дифференциальный рецептор). Поведение тепловых рецепторов при изменениях температуры как в зеркале отражает поведение холодовых: при нагревании их импульсация растет выше стационарного уровня, а при охлаждении падает ниже него, после чего возвращается к этому уровню. Температурные пределы динамической чувственности терморецепторов отличаются от установленных для статической. Например, при охлаждении кожи ниже максимума статической импульсации холодового рецептора динамическая частота его импульсации повышается, несмотря на решение ее статического уровня.
Следовательно, в целом выгоднее иметь и тепловые, и холодовые терморецепторы, а не дородную их популяцию, поскольку это дает ДНС значительно более детальную информацию о термических процессах в коже.
2.5 Висцеральная чувствительность
Спинномозговые и некоторые черепномозговые нервы содержат афференты, идущие от внутренних органов. Главные периферические рвы, в которых они проходят, - блуждающий, внутренностньй и тазовый. Эфферентные волокна их нервов относятся к парасимпатической и симпатической системам, а афферентные – к другой.
Информация, проводимая висцеральными афферентами, используется главным образом для вегетативных регуляторних процессов, управляющих кровообращением, дыханием, пищеварением, электролитическим и водным балансами.
Активность висцеральных афферентов до которой степени воспринимается сознательно. Это могут быть ощущения, обусловленные непосредственно данными волокнами или же вызванные опосредованно рефлексами с их участием. По логике вещей висцеральные афференты действуют на наше сознание тем сильнее, чем больше мы способны противодействовать регуляторным нарушениям сознательными поведенческими реакциями. Сильная, потенциально вредная стимуляция внутренних органов воспринимается как боль; при недостаточном потреблении пищи или воды активность висцеральных афферентов (например, глюкозных рецепторов или осморецепторов) вызывает «общие» ощущения голода или жажды; заполнение мочевого пузыря в конце концов побуждает нас его опорожнить. В диапазоне между этими конкретными восприятиями и полностью бессознательными регуляторными реакциями висцеральные афференты вызывают также неспецнфичные приятные или неприятные ощущения, в общем определяемые как самочувствие и обычно сильно влияющие на эмоциональное состояние.
Сердечно-сосудистая система
В регуляции давления и объема крови участвуют механорецепторы в аорте, сонной артерии и предсердиях. Их непрерывную активность мы не осознаем. Однако в некоторых особых ситуациях нами ощущается собственная сердечная деятельность, например при сильном физическом или психическом напряжении. В этом восприятии сердцебиений, вероятно, также участвуют механорецепторы в коже и мышцах, возбуждаемые значительными изменениями формы и положения сердца, а также пульсацией артерий. Речь идет о непрямом восприятии активности висцеральных афферентов через запускаемые ими рефлексы.
Легочная система
Афференты, регулирующие дыхание, - это прежде всего хеморецепторы в артериальной системе и, возможно, также в мозге, возбуждающиеся понижением парциального давления О2 и повышением его у СО2. Активность этих рецепторов тоже, как правило, не ощущается. Только когда парциальное давление СО2 резко возрастает и/или воздухоносные пути перекрываются (например, при астме), человек чувствует, как задыхается. Снижение парциального давления О2 из-за недостатка кислорода во вдыхаемом воздухе (например, при восхождения на высокую гору или при падении давления в кабине самолета) не ощущается, даже если оно настолько сильно, что вызывает полубессознательное состояние. Следовательно, непосредственное участие хеморецепторов, анализирующих кровоток, в появлении одышки маловероятно; воспринимается скорее недостаток двигательного выполнения дыхательной программы. Ритмичные дыхательные движения осознаются, вероятно, благодаря возбуждению многих механорецепторов в грудном скелетно-мышечном аппарате и коже.
Желудочно-кишечная система
По своему эмбриологическому происхождению желудочно-кишечный тракт-часть поверхности тела. Их функциональное соответствие заключается в том, что при переработке проглоченной пищи на желудок и кишки действует множество стимулов, возникающих во внешней среде. Как и следовало ожидать, механические, тепловые и химические стимулы воспринимаются желудочно-кишечной системой сильнее, чем другими внутренними органами. Особенно отчетливо она реагирует на растяжение; это можно доказать экспериментальным раздуванием баллона, введенного в пищеварительный тракт при помощи катетера. Растяжение желудка вызывает чувство наполнения, или сытости, растяжение кишечника – чувство закупорки (как при кишечных газах). В таких опытах с надуваемым баллоном стимулы иногда ошибочно локализуются испытуемым в зонах Геда на поверхности кожи. Сильная стимуляция желудочно-кишечной системы растяжением ощущается как боль, особенно когда вызывает контрактуру гладкой мускулатуры.
Тепловые и химические стимулы воспринимаются главным образом в пищеводе и прямой кишке. Судя по наблюдениям, сделанным в то время, когда операции на органах брюшной полости производились без наркоза, прикосновение к желудочно-кишечным структурам и их пальпация не осознаются.
Почечная система
В области почек и мочеточников человек способен ощущать только боль. Однако, если мочевой пузырь настолько наполнен, что растягивается, возникает потребность его опорожнить, обусловливающая произвольное мочеиспускание.
2.6 Проприоцепция
Восприятие позы и движения нашего собственного тела называется проприоцепцией (а также глубокой, или кинестетической, чувствительностью). Поза определяется углом расположения костей в каждом суставе, устанавливаемым либо пассивно (внешними силами), либо активно (мышечным сокращением). Рецепторы проприоцепции находятся не только в суставах, но также в мышцах сухожилиях и коже. С их работой сочетаются сигналы от вестибулярного органа, что позволяет определять положение тела в поле земного притяжения. Все эти проприоцепторы участвуют также и в нашей сознательной и бессознательной двигательной активности.
Типы проприоцепции
Чувство позы. Даже с закрытыми глазами мы осознаем углы между костями в наших суставе а значит, и относительное положение частей тела. Эту способность легко продемонстрировать на простых опытах, выполняемых испытуемыми с закрытыми глазами. Если конечности на одной стороне тела пассивно или активно придано какое-нибудь положение (например, рука согнута в локте), испытуемый может точно воспроизвести его конечностью на другой стороне. Если попросить его показать на себе определенный участок тела, он сможет это сделать с очень небольшой ошибкой.
Чувство движения. При изменении положения сустава мы воспринимаем и направление, и скорость этого движения. Амплитудный порог такого восприятия зависит от угловой скорости.
Чувство силы. Благодаря ему мы воспринимаем степень мышечного усилия, нужного для выполнения движения или поддержания позы (например, для разных гравитационных нагрузках), и достаточно хорошо оцениваем вес предметов, поднимая их рукой. Сравнивая вес двух предметов, одновременно поднимаемых обеими руками, мы воспринимаем разницу уже на уровне 3-10%. Если сравниваемые предметы положить на руки, а те на подставку, оценки веса, основанные теперь на афферентной информации от кожных рецепторов, будут значительно менее точны.
Проприоцепторы
Сознательное восприятие движения и положения суставов, по-видимому, обусловлено одновременным действием различных механорецепторов: суставных и кожных рецепторов, мышечных веретен, сухожильных органов. С этим согласуется и то, что соматосенсорное ядро таламуса (вентробазальный юмплекс) и соматосенсорная кора получают информацию от всех перечислении выше их типов. В последние годы возникли некоторые разногласия по поводу относительного участия в восприятии каждого из них.
Долго считалось, что проприоцепция обусловлена главным образом суставными рецепторами. Эти механорецепторы в суставных капсулах кодируют различные положения или движения суставов. Однако их предполагаемая особая роль в проприоцепции стала оспариваться, когда было установлено, что больные с искусственными суставами различает их положение почти так же, как обычные люди. Более поздние психофизические опыты по восприятию положения и движения суставов подтвердили гипотезу, согласно которой важнейшую роль в нем играют мышечные веретена. Воздействуя на мышцы и сухожилия вибрацией, активирующей главным образом мышечные веретена и сухожильные органы, можно вызывать иллюзии движения в суставах, сбивающие испытуемых с толку.
Вывод, вытекающий из многочисленных психофизических исследований и клинических наблюдений, состоит в том, что для проприоцепции ЦНС использует всю доступную ей нервную информацию. По всей вероятности, афферентные сигналы от мышечных веретен, сухожильных органов, суставных и кожных рецепторов интегрируются, а это приводит к сложному восприятию положения и движения тела. Эфферентная двигательная информация, например команды от двигательной коры, скорее всего, также используется в этом процессе в виде копии эфферентации.
2.7 Функциональный и анатомический обзор центральной соматосенсорной системы
Центральную переработку сигналов от периферических рецепторов можно рассматривать на трех функциональных уровнях - афферентной, интегративной и эфферентной подсистем. Афферентный, или сенсорный, уровень соматовисцеральной системы связан со спинным мозгом, стволом мозга, таламусом и корой больших полушарий. Действуют различные проводящие пути, позволяющие перерабатывать информацию параллельно, используя ее для выполнения разных функций. Периферическая сенсорная поверхность (совокупность всех рецепторов вне ЦНС) проецируется на таламус и кору таким образом, что упорядочение (соматотопически) картируется в ЦНС. Следующие этапы центральной нервной переработки информации соответствуют интегративному и эфферентному уровням. Ассоциативная и лимбическая системы выполняют главным образом интегративные функции. Одна из важнейших задач этих систем—сопряжение сигналов от различных сенсорных систем с информацией, хранящейся в памяти. Двигательная и вегетативная системы выполняют эфферентные функции. Сложные взаимодействия трех функциональных подсистем, по-видимому, лежат в основе поведения. Оно в присутствии сенсорных стимулов делится на восприятия и реакции с пятью компонентами: когнитивным (включающим осознанное различение), аффективным (связанным с эмоциями), мотивационным (связанным с побуждениями, или драйвами), двигательным и вегетативным.
Дихотомия в восходящей центральной соматовисцеральной системе Афферентные нервные волокна от туловища и конечностей идут в спинной мозг по спинномозговым нервам, а от лица – в ствол мозга по тройничному нерву. Сигналы от рецепторов различным образом используются в обоих этих центральных отделах для двигательных и вегетативных рефлексов, а также передаются в высшие отделы головного мозга по системам восходящих волокон. На рисунке 2.3 для общей ориентации в таких связях дана сильно упрощенная их схема.
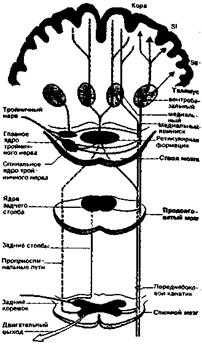
Рис. 2.3. Анатомическая организация главных функциональных элементов соматосенсорной системы (схема).
Соматосенсорная информация передается по двум главным системам восходящих трактов, различающихся как анатомически, так и функционально. Это системы:
- заднего столба, или лемнисковая;
- переднебокового канатика.
Главные связи в обеих системах служат для соединения каждой стороны тела с противоположной (контралатеральной) половиной головного мозга. Функционально система заднего столба связана с кожной механорецепцией и проприоцепцией, а система переднебокового канатика – главным образом с терморецепцией и ноцицепцией. Хотя в спинном мозгу обнаружены дополнительные восходящие пути соматосенсорной информации, полезно сохранять именно такое деление, основанное на двух главных системах трактов.
Последствия повреждения спинного мозга
Когда спинной мозг только «надорван» с одной стороны (например, при несчастном случае), прерывание трактов в его белом веществе вызывает характерный неврологический дефицит, синдром Броун-Секара. Он заключается в сенсорных нарушениях с обеих сторон тела ниже уровня повреждения с параличом произвольных движений на ипсилатеральной стороне. Сенсорная недостаточность с двух сторон тела неодинакова: испилатерально нарушено осязание (повышен, например, двухточечный порог, а контралатерально-восприятие боли и тепловых стимулов. В то же время ипсилатеральные ноцицепция и терморецепция, как и контралатеральное осязание, нормальные. Поэтому такой эффект называется диссоциированным сенсорным дефицитом.
Он обусловлен разрывом трактов в белом веществе. Блокада нисходящих двигательных путей (например, пирамидного тракта) вызывает ипсилатеральный паралич произвольных движений, повреждение заднего столба нарушает ипсилатеральное осязание, а переднебокового канатика – контралатеральные ноцицепцию и терморецепцию. Сенсорная информация от определенной области кожи идет по заднему столбу и переднему канатику с обоих сторон, но на уровне головного мозга в случае обоих этих сенсорных путей она воспринимается преимущественно контралатерально той стороне тела, на которой действует стимул.
Система заднего столба
Название этой системы отражает тот факт, что сенсорные сигналы передаются по заднему столбу (или канату), т.е. массивному пучку волокон в дорсальном белом веществе спинного мозга. Он состоит из прямых коллатералей толстых миелинизированных афферентов спинномозговых нервов. Другое название – лемнисковая система – обусловлено тем, что выше спинного мозга ее сигналы проводятся по медиальному лемниску (медиальной петле). Эта восходящая соматосенсорная подсистема соединяет спинномозговые и тройничные механорецептивные афференты с двумя анатомически и нейрофизиологически разделенными участками теменной коры – первой и второй соматосенсорными областями.
Афференты в системе заднего столба идут от низкопороговых кожных механорецепторов (МА, БА и ТП), мышечных веретен, сухожильных органов и суставных рецепторов. Проведение в этом восходящем пути быстрое, поскольку здесь только три синаптических переключения. Он состоит из следующих основных частей (рис. 2.3): задних столбов спинного мозга, ядер заднего столба в продолговатом мозгу (первый синапс), медиального лемниска, перекреста (переход путей на другую сторону), вентробазалъного ядра таламуса (второй синапс), корковых областей SI и SII (третий синапс). Тройничные коллатерали образуют синапсы в главном тройничном ядре ствола мозга, соответствующем ядрам заднего столба. После перекреста проводящий путь продолжается по медиальному лемниску к таламусу.
Особое свойство системы заднего столба – упомянутая выше соматотопическая организация, т.е. примерное сохранение топографических связей разных областей кожи (периферической сенсорной поверхности) во всех центральных зонах переключения. Это приводит к геометрически искаженному картированию, или проецированию, первых на вторые, подобные тому, которое свойственно зрительной и слуховой системам.
Система заднего столба особенно хорошо развита у приматов, т.е. у обезьян и человека. Она представляет собой анатомический субстрат тактильных ощущений (осязания) и проприоцепции, всех сознательных и бессознательных способностей, требующих различения пространственных и временных деталей механической стимуляции тела.
Сигналы от кожи, мышц и суставов, передаваемые по заднему столбу (особенно тактильные), дают важную информацию о выполнении движений. Поэтому у больных с повреждением заднего столба сильно понижена способность распознавать предметы, например, активно ощупывая их, или идентифицировать цифры, вычерчиваемые на коже. Двухточечный порог у них повышен.
Система переднебокового канатика
Эта система показана на рис. 2.3. Переднебоковой канатик включает в себя ряд трактов в вентральном белом веществе спинного мозга, содержащих восходящие волокна спинальных нейронов, тела которых находятся в сером веществе, главным образом в заднем роге. Тройничные аксоны, соответствующие переднебоковому канатику, идут от нейронов в спинальном тройничном ядре (рис. 2.3). Афферентные сигналы в переднебоковой канатик проводятся спинномозговыми и тройничными нервами от терморецепторов и ноцицепторов, а также от низкопороговых механорецепторов, в основном от расположенных в коже. Восходящие аксоны присоединяются к переднебоковому тракту, контралатеральному телам их клеток и входам их афферентных волокон в спинной мозг. Поскольку главных мест назначения два - ретикулярная формация с другими стволовыми областями мозга и таламус, различают спиноретикулярный и спиноталамический тракты. Передаваемая ими информация, по-видимому, в конце концов достигает многих отделов головного мозга, главным образом по полисинаптическим, медленно проводящим путям. Здесь нет ни четкой соматотопической организации, ни ясно очерченной проекции на кору, сравнимой с известной для системы заднего столба. Система переднебокового канатика рассматривается как анатомический субстрат терморецепции и ноцицепции и, кроме того, как часть неспецифичной системы.
Неспецифичная система
Приблизительно с 1950 г. в нейрофизиологии распространилось понятие неспецифичной системы как альтернативы четко определимым анатомически и физиологически специфичным системам различных сенсорных модальностей. В соматосенсорной модальности система заднего столба – специфичная. Неспецифичную систему рассматривали как субстрат ряда наблюдаемых в опытах на животных генерализованных peaкций типа:
- реакций возбуждения;
- изменения ЭЭГ;
- ритма сон/бодрствование;
- аффективных поведенческих реакций;
- изменений кровяного давления.
Они могут быть вызваны как сенсорными стимулами (особенно болевыми), так и электрической стимуляцией ретикулярной формации ствола мозга и медиальных таламических областей. Эти центральные области считались «сердцевиной» неспецифичной системы, получающей сенсорную информацию главным образом по переднебоковым трактам.
Исследования названного выше филогенетически древнего отдела головного мозга показывают, что здесь протекают многие жизненно важные интегративные и регуляторные процессы, отражающиеся на функциях спинного (например, симпатические реакции) и большого мозга (например, состояние тревоги). Однако достаточных свидетельств существования однородной неспецифичной системы нет. Напротив, с накоплением данных по физиологической, анатомической и биохимической дифферецировке этих отделов головного мозга сам такой термин становится все менее и менее приемлемым.
2.8 Передача соматовисцеральной информации в спинном мозгу
Дерматомы
Афференты от кожи, мышц, cycтавов и внутренних органов входят в спинной мозг задним корешкам в определенном пространственном порядке, т. е. согласно топологической организации. Кожные афференты каждого заднего корешка иннервируют ограниченную область кожи, называемую дерматомом. Смежные дерматомы сильно перекрываются из-за перераспределения пучков волокон при их распространении к периферии, особенно в плечевом и пояснично-крестцовом сплетениях. Один периферический нерв содержит волокна нескольких соседних задних корешков, а каждый задний корешок – волокна разных нервавов. В то время как перерезка периферического нерва вызывает сенсорный дефицит ограниченного участка тела, перерезка одного заднего корешка, хотя и обедняет иннервацию какой-то области, все же не вызывает в ней заметного сенсорного дефицита.
В скелетной мускулатуре эквивалентами дерматомов являются миотомы. Зона Геда внутреннего органа состоит из дерматомов, связанных со спинномозговыми сегментами, иннервирующими этот орган. Нейронная организация обеих совокупностей афферентов такова, что может вызывать отраженную боль.
Функциональные свойства спинальной соматовисцеральной системы
Соматовисцеральные афференты образуют синаптические связи с нейронами в заднем (дорсальном) роге спинного мозга. Некоторые толстые миелинизированные афференты (группы I ІI) также отделяют ветви (коллатерали), восходящие в составе спинного мозга. У заднего рога, где идет нейронная переработка информации, четыре выхода: длинные тракты, восходящие до головного мозга главным образом в переднебоковом канатике; восходящие и нисходящие проприоспинальные связи с соседними сегментами; сегментарные связи с мотонейронами и симпатическими нейронами (участие в спинальных двигательных и вегетативных рефлексах). Однако, по всей вероятности, один нейрон не образует все четыре вида связей сразу.
В восходящих трактах аксоны, идущие от одного сегмента, расположены бок о бок. Это создает соматотопическую слоистость; аксоны, присоединяющиеся к прочим во все более высоких сегментах, всегда оказываются со стороны серого вещества. Один из результатов такого расположения состоит в том, что при поверхностном повреждении переднебокового канатика (раной или опухолью) в шейной области первые сенсорные дефициты следует ожидать в нижней половине тела.
Нейроны переднебокового канатика
Хотя клинические данные указывают на участие переднебокового канатика в болевых и температурных ощущениях, он содержит всего лишь несколько аксонов специфичных ноцицептивных и терморецептивных нейронов. Большинство аксонов принадлежит здесь клеткам, возбуждаемым также слабыми механическими стимулами, т.е. мультирецептивным нейронам. Остается спорным вопрос, достаточно ли немногих специфичных ноцицептивных и терморецептивных нейронов для передачи соответствующих сигналов или дополнительная информация, требуемая головному мозгу для болевых и температурных ощущений, обеспечивается активностью мультирецептивных нейронов. Важное свойство заднего корешка как станции переключения – конвергенция афферентов от кожи и внутренних органов на одни и те же нейроны. Из-за нее афферентные сигналы от внутренностей (например, запускаемые желчными коликами) «ложно» локализуются больным на поверхности тела. Каждый внутренний орган связан с определенной областью кожи – своей зоной Геда. Другой элемент цепи заднего корешка – тормозные синапсы, которые служат для модификации афферентной информации. Это может принимать форму латерального торможения или регуляции сенсорного входа трактами, нисходящими от головного мозга. Активация таких тормозных механизмов применима для обезболивания.
Нейроны ядер заднего столба
Аксоны заднего столба оканчиваются ипсилатерально в его ядрах (клиновидном и тонком), находящихся в продолговатом мозгу. Здесь они образуют синапсы с крупными нейронами, посылающими аксоны в контралатеральный таламус по медиальному лемнисовому тракту и называемыми релейными в отличии от вставочных (интернейронов) с аксонами выходящими из ядер заднего столба. Переработка афферентной информации при синаптической передаче к релейным нейронам имеет следующие характеристики:
- сохранение сенсорной специфики; на одном нейроне конвергируют афференты одинакового сенсорного типа;
- большой запас надежности синаптической передачи; даже одиночные импульсы aфферентного волокна вызывают постсинаптическую импульсацию;
- мелкие рецептивные поля;
- соматотопическая организация; пространственно упорядоченное представители кожи;
- афферентное торможение;
- нисходящие регулирующие входы, в частности, от коры головного мозга.
Эти свойства характерны и для последующих мест переключения в лемнисковой системе; они определяют качество и разрешающую способность ощущений, связанных с передаваемой здесь информацией.
2.9 Соматосенсорные функции ствола мозга
Ствол мозга, состоящий из продолговат мозга, моста и среднего мозга представляет собой место отхождения (или вхождения) большинства черепно-мозговых нервов. В нем содержится множество дискретных ядер, выполняющих преимущественно сенсорные, двигательные или регуляторные функции; между ними, по-видимому, проходят все восходящие и нисходящие пути связывающие головной мозг со спинным, а также большой мозг с мозжечком.
Тройничный нерв и его центральные связи
Голова иннервируется 12 парами черепно-мозговых нервов, большинство которых присоединяется к центральной нервной системе в стволе мозга и промежуточном мозгу. Пятый из этих нервов называемый тройничным из-за своих трех ветвей, включает афференты от лица и ротовой области (рис. 2.4).
Он иннервирует кожу, зубы, слизистую рта, язык и роговицу. Седьмой черепномозговой нерв (лицевой) тоже, но в меньшей степени, содержит головные соматосенсорные афференты. В языкоглоточном и блуждающем нервах есть висцеральные афференты от органов кровообращения, дыхания и пищеварения.
Сенсорные тройничные ядра и восходящие пути
Афференты тройничного нерва образуют синапсы в ядре спинального тракта и главном сенсорном ядре (рис. 2.4). Ядро спинального тракта функционально соответствует заднему рогу спинного мозга. Здесь механорецептивная, терморецептивная и ноцицептивная информация передается на аксоны, несущие ее к ретикулярной формации и таламусу, как и волокна переднебокового канатика, передающие туда информацию от спинного мозга. Все афференты, которые оканчиваются в главном сенсорном ядре, соответствующем ядрам заднего столба, идут от низкопороговых механорецепторов. Постсинаптические аксоны переходят на другую сторону и направляются к таламусу в составе медиального лемнискового тракта.
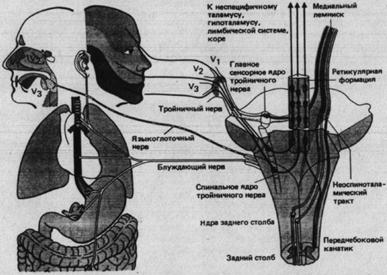
Рис. 2.4. Афференты и
структуры соматовисцеральной системы в стволе головного мозга (справа - его
схематичный вид сзади). К ним относятся афференты тройничного нерва (трех его
ветвей V![]() , V
, V![]() и V
и V![]() ), блуждающего и языкоглоточного
нервов, центральная афферентная система тройничного нерва и ее восходящие пути,
ретикулярная формация и тракты, приходящие из спинного мозга, т.е. дорсальных
столбов и переднебокового канатик. Центральное прохождение афферентов
языкоглоточного и блуждающего нервов не показано.
), блуждающего и языкоглоточного
нервов, центральная афферентная система тройничного нерва и ее восходящие пути,
ретикулярная формация и тракты, приходящие из спинного мозга, т.е. дорсальных
столбов и переднебокового канатик. Центральное прохождение афферентов
языкоглоточного и блуждающего нервов не показано.
В стволе мозга информация, приносимая тройничными афферентами, интегрируется в двигательные рефлексы мускулатуры головы и во множество вегетативных рефлексов. Тройничная система осуществляет жизненно важные функции (особенно у млекопитающих) тактильного исследования окружающей среды, питания, издавания звуков и т.д. Она хорошо развита уже при рождении и проводит сенсорные сигналы, запускающие пищевое поведение, т.е. обеспечивает первое сенсорное знакомство ребенка с окружающей средой.
Ретикулярная формация
Ретикулярная формация – протяженная структура в стволе мозга (рис. 2.4)-важная интегративная область неспецифичной системы. Соматовисцеральные афференты попадают по спиноретикулярному тракту переднебокового канатика, а возможно, также по проприоспинальным (полисинаптическим) путям и соответствующим путям от ядра спинномозгового тройничного тракта. У ретикулярной формации есть афферентный вход и от других черепно-мозговых нервов. Кроме того, здесь много различных эфферентных связей: нисходящих к спинному мозгу и восходящих через неспецифичные таламические ядра к коре больших полушарий, гипоталамусу и лимбической системе.
Функции ретикулярной формации еще не вполне понятны. Ретикулярная формация участвует в:
- регуляции возбудимости коры: уровня осознания стимулов и реакций, ритма сон/ бодрствование (восходящая активирующая ретикулярная система);
- придании аффективно-эмоциональных аспектов сенсорным стимулам, особенно болевым, за счет передачи афферентной информации в лимбическую систему;
- двигательной регуляции, особенно связанной с так называемыми жизненно важными рефлексами (кровообращения, дыхания, глотания, кашля и чихания), требующими координации нескольких афферентных и эфферентных систем;
- регуляции позы и целенаправленных движений.
Этот перечень свидетельствует о невозможности четкого различения в ретикулярной формации цента сенсорной и интегративной систем. С другой стороны, недавно стало ясно, что некоторые ее области можно различать по анатомическим, функциональным и химическим свойствам (например, серотонинергических ядра шва и норадренергическое голубое пятно). Скорее всего, по мере расширения наших знаний о ретикулярной формации, представление о неспецифичной системе будет пересмотрено.
2.10 Таламус
Таламус считается входными воротами и распределительным пунктом, через которые все афферентные системы получают доступ к филогенетически более молодым церебральным структурам, обеспечивающим осознание сенсорных стимулов и сознательное целенаправленное поведение.
Анатомический и функциональный обзор
Правый таламус можно разделить на несколько функционально и/или анатомически различимых ядер, каждое из которых связано со своей корковой областью. Для обобщей ориентации сгруппируем эти ядра в следующие четыре функциональных класса:
- специфичные переключающие и перерабатывающие ядра кожных сенсорных органов, глаза и уха;
-ядра с преимущественно двигательными функциями;
-ядра с ассоциативными функциями;
- неспецифичные ядра без определенных корковых мишеней.
Таламические переключающие ядра сенсорных органов
Эти переключающие и перерабатывающие структуры связаны с корковой областью, отвечающей за их сенсорную модальность, и в свою очередь регулируются (возбуждаются и тормозятся) этой областью.
Неспецифичные ядра
Эта категория включает медиальные области, примыкающие к третьему желудочку мозга, а также интраламинарные ядра. Это высшие перерабатывающие и распределительные пункты для афферентной информации, конвертирующей на ретикулярной формации мозгового ствола. Сигналы от спинного мозга приходят сюда непосредственно по палеоспиноталамическому тракту и непрямо – по спиноретикулярному.
Двигательные ядра
Самое главное из них вентролатеральное (ВЛ), связывающее мозжечок и базальные ганглии с двигательной корой. Хирургические воздействия на соответствующие участки ВЛ могут ослабить двигательные расстройства (например, паркинсонизм).
Ассоциативные ядра
Эти части таламуса соединены с корой, но не могут быть отнесены к какой-либо определенной сенсорной системе; они принимают участие в интегратнвных функциях головного мозга.
Специфичное таламическое ядро соматосенсорной системы
Из-за своего анатомического положения этот участок лемнисковой системы называется вентробазальным ядром (ВБ) или вентробазалъным комплексом. В нем выделяют вентральное постеролатеральное (ВПЛ) и вентральное постеромедиальное (ВПМ) ядра. В ВПЛ находится нейронное представительство туловища и конечностей, а в ВПМ — лица. Главный проводящий путь, который ведет к ВПЛ, - это медиальный лемнисковый тракт, а к ВПМ тройнично-таламический тракт, начинающийся от главного сенсорного ядра тройничного нерва.
В опытах на наркотизированных животных установлены следующие функциональные особенности ВБ-нейронов:
-каждый из них обладает определенным рецептивным полем для механических стимулов кожи;
- рецептивные поля тем меньше, чем дистальнее они находятся на конечностях;
- соседние участки тела проецируются на соседние участки ВБ (пример соматотопической организации);
- каждый нейрон возбуждается главным образом рецептором одного типа (например, рецепторами МА или волосяных фолликулов в коже);
- импульсация нейрона усиливается с ростом интенсивности периферической стимуляции, т.е. интенсивность кодируется так же, как у кожных механорецепторов.
2.11 Соматосенсорные проекционные области в коре
Вентробазальный комплекс таламуса соединен как восходящими, так и нисходящими аксонами с двумя корковыми зонами – SI и SII (S означает «соматосенсорная область»). SI расположена на постцентральной извилине непосредственно позади глубокой центральной борозды, проходящей поперек полушария. SII лежит на верхней стенке боковой борозды, разделяющей теменную и височную доли. SI филогенетически моложе SII и очень важна у высших млекопитающих (особенно у приматов); она участвует во всех функциях соматосенсорной системы, зависящих от хорошего пространственно-временного различения стимулов. Некоторые из этих функций можно исследовать, например по методу двухточечного порога кожи (рис. 2.5)
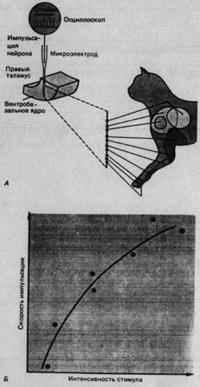
Рис. 2.5. Рецептивные поля нейронов в вентробазальном ядре таламуса. Микроэлектрод, проводимый через таламус наркотизированной кошки, прошел 10 нейронов, отвечающих на механическую стимуляцию кожи. Их рецептивные поля находились, как показано, на левой передней конечности.
Топографическая организация соматосенсорной коры
Соматотошпеское представительство периферии тела находится в ковтралатеральной SI, организовано поразительно подробно и было тщательно изучено. Вся поверхность тела здесь картирована причем у приматов даже имеются множествен проекции кисти. Сходное, хотя и менее четкое, картирование свойственно и SII, где оно частично билатерально. Соматотопическую opганизацию сенсорной коры изучают несколькими методами. У человека во время операции на головном мозгу применяют ее локальную электрическую стимуляцию с целью вызывать ощущения локализованные в соответствующих участках тела. Можно также с помощью радиоактивного ксенона измерять локальный корковый кровоток во время стимуляции кожи. У животных для картирования коркового представительства периферии тела использовали анализ вызванных потенциалов и запись активности одиночных нейронов.
Электрическая стимуляция коры мозга человека
При ряде нейрохирургических операций проводят локальную электрическую стимуляцию коры бодрствующего больного (для анестезии места разреза). Вызываемые этим ощущения воспринимаются так, как будто возникают на периферии. Систематическое обследование таким способом SI позволило получить картину, приведенную на рис. 2.6.
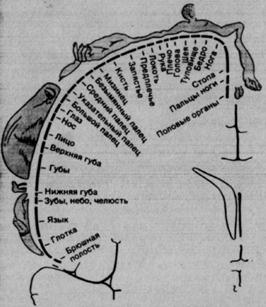
Рис. 2.6. Соматотопическая организация корковой зоны SI человека. Изображения над поперечным срезом мозга (на уровне постцентральной извилины) и их обозначения демонстрируют пространственное представительство поверхности тела в коре, установленное путем локальной электрической стимуляции мозга бодрствующих больных.
Этот «соматосенсорный гомункулус» – сильно искаженная карта периферической сенсорной поверхности с непропорционально крупными представительствами областей пальцев и pта. У человека они особенно густо иннервированы и, как показали психофизические опыты, характеризуются прекрасной пространственной разрешающей способностью, т. е. низкими двухточечными порогами. Очевидно, эти два факта причинно связаны, причем такая связь существует и в случае других сенсорных систем: чем больше рецепторов и центральных нейронов приходится на 1 мм2 сенсорной поверхности, тем лучше пространственное разрешение ею стимулов.
Переработка информации в нейронах соматосенсорной коры
Колонки корковых нейронов. При прохождении микроэлектрода перпендикулярно поверхности постцентральной извилины последовательно встречаемые им нейроны часто обладают идентичными или широко перекрывающимися рецептивными полями. Если же электрод перемещается под углом к этой поверхности, он проходит нейроны с соседними, но четко разграниченными рецептивными полями (в соответствии с соматотопической организацией). Из этих и других данных сделан вывод, что соматосенсорная кора, как и двигательная, организована в виде функциональных единиц-колонок нейронов, перпендикулярных ее поверхности.
Рецепторная специфичность колонок
Адекватная избирательная стимуляция, например, различных кожных рецепторов показывает, что нейроны одной колонки возбуждаются обычно рецепторами только одного типа. Очевидно, колонки – это функциональные единицы, соответствующие положению и сенсорной специфике периферических сенсорных нервных окончаний.
Простые и сложные нейроны
Характеристики импульсации простых корковых нейронов очень близки к таковым у связанных с ними рецепторов. Например, в SI есть нейроны, ведущие себя как рецепторы БА. Корковые нейроны, отвечающие на периферические стимулы импульсацией, явно отличающейся от той, что свойственна связанным с ними рецепторам, называются сложными. Этот термин охватывает разнообразные клетки. Среди них известны нейроны, которые реагируют только на стимулы, движущиеся линейно по поверхности кожи, и дают максимальный ответ на движение в определенном направлении. Они обнаружены в SI, SII и ассоциативных областях теменной коры.
Терморецептивные корковые нейроны, найденные у обезьяны, относятся к сложным. В отличие от периферических терморецепторов они реагируют либо только на изменения температур кожи, либо только на ее постоянную температуру. Кроме того, у них нет пика ответа при температуре около 26 оС, характерного для холодовых рецепторов. Другой тип сложных терморецепторных нейронов обнаружен в коре мозга крысы. Здесь частота импульсации «скачет» между максимумом и минимумом, когда температура соответствующих рецептивных полей кожи меняется приблизительно на 2 оС (пороговый детектор).
Эти примеры простых и сложных нейронов показывают, что корковая переработка сигналов приводит к нейронному представительству или отфильтровыванию информации о самых разнообразных параметрах периферических стимулов. Такая способность извлекать информацию, названная выделением признаков, особенно тщательно изучена в зрительной коре.
Эфферентные связи SI
От области SI, как и от всех прочих участков коры, отходит множество эфферентных аксонов. Как полагают они проводят в переработанном виде информации о периферических стимулах к другим частям центральной нервной системы. SI связана со следующими областями (указана и основная функция связи):
-двигательная кора: регуляция движения по обратной связи;
-теменные ассоциативные зоны: интеграция тельной и тактильной информации;
- контралатералъные SI и SII: интеграция билатеральной тактильной информации;
- таламус, ядра заднего столба, спинной мозг: эфферентная регуляция потока афферентной информации.
Корковая область SII значительно меньше SI. Она лежит у латерального конца постцентральной извилины, на верхней стенке сильвиевой борозды. Здесь соматотопическое представительство поверхности тела билатеральное. Рецептивные поля нейронных колонок обычно находятся на обеих сторонах тела, а их проекции часто расположены симметрично. Как полагают, SII играет особую роль в сенсорной и двигательной координации двух сторон тела (например, при хватании или ощупывании обеими руками).
Соматосенсорная кора и восприятие
Многие данные говорят о том, что SI необходима для точного тактильного различения и сознательного восприятия событий на поверхности и с высоким разрешением их силовых, пространственных и временных деталей. Впрочем, возбуждений SI необязательно осознается; например, периферическая стимуляция дает первичные вызванные потенциалы у наркотизированного или спящего человека сознание которого отключено.
Стимуляция коры человека
Локальная электрическая стимуляция SI или SII бодрствующего больного вызывает у него ощущения, которые часто идентичны возникающим при соответствующей стимуляции на периферии. Сенсорные ощущения возникают также (и только) при раздражении SII, зрительной и слуховой проекционных зон. Действие околопороговых точечных стимулов на SI описывалось больными, сходное с «естественной стимуляцией». При электрическом воздействии на кору возникали простые ощущения, специфичные для определенных рецепторов (вибрация, тепло, холод), ощущение перемещающихся по коже стимулов или движений суставов. О болевых ощущениях пациенты сообщали редко.
Удаление коры
Когда часть коры в области SI человека повреждена или удалена хирургическим путем, возникает дефицит восприятия. Стимуляция кожи еще может быть воспринята как таковая, но способность различать пространственные детали раздражителя снижена. Степень дефицита зависит от размеров поврежденного участка коры и с течением времени постепенно уменьшается. Как полагают, это улучшение наступает вследствие способности других корковых областей принимать на себя функции.
Корреляции между соматосенсорной нейрофизиологией и психофизикой
В последнее время исследовались количественное соотношение между восприятием и уровнем активации одиночных нейронов или популяций. Полученные результаты часто позволяют делать выводы о нейронном субстрате восприятия.
В одном из опытов был применен электрический вибратор, вызывавший на кисти обезьяны синусоидальную механическую деформацию кожи разной частоты и амплитуды. Порог обнаружения (восприятия) этого стимула определяли по инструментальному условному рефлексу. Предварительно обезьянам вживляли под наркозом микроэлектрод, позволяющий одновременно регистрировать электрофизиологическую активность нейронов в SI, в частности измерять порог в реакции на механический стимул. Была установлена количественная корреляция между обоими порогами. Один из выводов, какой можно извлечь из этого опыта: данные нейроны SI участвуют в процессах восприятия действующих на кожу механических стимулов.
2.12 Контроль афферентного входа в соматосенсорной системе
На всех уровнях ЦНС афферентная информация может испытывать различные влияния и изменяться (модулироваться). Это вызывается либо взаимным торможением афферентами друг друга (афферентным торможением), либо центробежным (нисходящим) торможением. Несколько примеров афферентного торможения уже рассматривались: это аутогенное и антагонистическое торможение в спинальных двигательных системах и латеральное торможение во многих сенсорных системах.
Центробежная регуляции афферентных сигналов
Сенсорные системы нельзя рассматривать как улицы с односторонним движением, по которым информация передается от периферии к коре; напротив, на промежуточную переработку такой информации влияет целый ряд (возбуждающих или тормозных) сигналов центрального происхождения, проводимых в противоположном направлении. Схема на рис. 2.7, А показывает нисходящие тормозные пути соматосенсорной системы, начинающиеся в коре и стволе мозга.
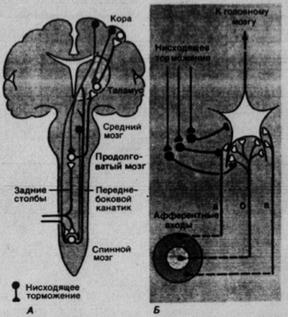
Рис. 2.7. Центробежная регуляция соматосенсорной системы. А. Афферентная информация может модулироваться в синапсах нисходящим торможением. Б. Пример функций нисходящего торможения. В синапсах, через которые активность афферентов (а, б, в) передается центральному соматосенсорному нейрону, оно может изменять величину рецептивного поля этого нейрона, если афференты, идущие от периферической части рецептивного поля (а, в), тормозятся сильнее, чем идущие от его центра (б)
Что обеспечивают эти системы нисходящего торможения? Их известные или предполагаемые функциональные влияния на афферентную информацию можно обобщить следующим образом:
- порог афферентной синаптической передачи повышается. Одно из следствий этого – подавление обыденной информации (например, непрерывной стимуляции одеждой);
- размеры рецептивного поля центрального нейрона уменьшаются при усилении нисходящего торможения (рис. 2.7, Б);
- изменяется модальность нейрона, на котором конвергируют разные типы афферентов (рис. 2.7, Б, а, б);
- регулируется чувствительность или устанавливаются пределы передачи афферентной информации.
Регуляция чувствительности афферентной передачи нисходящим торможением
Рис. 2.8. Торможение соматосенсорной информации в спинном мозгу стимуляцией среднего мозга. А. Схема нисходящей системы и экспериментальной методики ее изучения. Стимулирующий электрод вживлен в центральное серое вещество (ЦСВ) среднего мозга наркотизированного животного. Другой электрод регистрирует здесь активность одиночных нейронов в ответ, например, на раздражение кожи очень сильным нагреванием. Б. Peaкция спинального нейрона на нагревание кожи до (слева) и во время (справа) электрической стимуляции ЦСВ. Зависимость между частотой импульсации спинального нейрона (ось ординат) и интенсивностью нагревав (температура кожи, ось абсцисс) до и во время стимуляции ЦСВ.
Рис. 2.8 показывает, что афферентная передача от кожных рецепторов может тормозиться в нейронах спинного мозга электрической стимуляцией среднего мозга. Тормозные стимулы идут по нисходящему пути и действуют либо пре-, либо постсинаптически. Тормозятся как ноцицептивные, так и неноцицептивные сигналы. В этом примере сенсорным стимулом служит температура кожи повреждающих уровней; ее уровень кодируется приблизительно линейно частотой импульсации спинальных нейронов (рис. 2.8, В). Стимуляция среднего мозга ее снижает (рис. 2.8, Б), из-за чего наклон кривой, отражающей кодирование интенсивности, уменьшается (рис. 2.8, В). Здесь нисходящее торможение можно рассматривать как механизм настройки чувствительности или изменения коэффициента усиления афферентной передачи; реакции на стимулы при любой интенсивности последних уменьшаются в одинаковое число раз в зависимости от степени нисходящего торможения.
О том, каким образом такие тормозные системы активируются в естественных условиях, известно еще мало. Одна из их функций, как полагают,- содействовать вниманию. Было высказано предположение, что иногда информация, восходящая от спинного мозга, может сама активировать нисходящее торможение. Это был бы случай возвратного торможения (торможения по обратной связи), отличающегося от спинального сегментарного возвратного (через клетки Реншоу) торможения с участием супраспинальной петли. В этом особом cлучае торможения по обратной связи его эффект может выражаться в автоматической установке пределов: наклон характеристической кривой (рис. 2.8, В), т. е. коэффицент усиления афферентной передачи, определяется интенсивностью ее самой. Такой механизм сравним, например с автоматической регуляцией коэффициента усиления во входе кассетного магнитофона.
Влияние двигательных систем на афферентную информацию
Двигательные центры тоже
могут оказывать значительное центробежное влияние на сигналы, поступающие от
рецепторов. Возьмем контроль состояния мышечного веретена системой ![]() -волокон, регуляцию
положения глаз глазодвигательной системой, ощупывающие движения пальцев при
тактильном исследовании и изменения мышечного тонуса в аппарате среднего уха.
Их считают механизмами центробежной модификации сенсорных каналов. Эти примеры
ясно показывают, что ЦНС участвует в восприятии, не только пассивно принимая
периферическую информацию, но и активно влияя на поток информации и регулируя
его множеством способов.
-волокон, регуляцию
положения глаз глазодвигательной системой, ощупывающие движения пальцев при
тактильном исследовании и изменения мышечного тонуса в аппарате среднего уха.
Их считают механизмами центробежной модификации сенсорных каналов. Эти примеры
ясно показывают, что ЦНС участвует в восприятии, не только пассивно принимая
периферическую информацию, но и активно влияя на поток информации и регулируя
его множеством способов.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Константинов А.И., Соколов В.А., Быков К.А. Сравнительная физиологическая сенсорная система. – М., 1980.
2. Серопегин И.М., Волков В.М., Синайский М.М. Физиология человека. – М., 1979.
3. Измайлов Ч.А. Психофизиология цветового зрения. – М., 1989.
4. Бертулис А.В., Глезер В.Д. Провтранственное цветовое зрение. – Л., 1990.
5. Шевелев И.А. Динамика зрительного сенсорного сигнала. – М., 1971.
6. Марр Д. Зрение. – М., 1987.
7. Дудкин К.Н. Зрительное восприятие и память. – Л., 1985.
8. Гуртовой Г.К. Глаз и зрение. – М., 1959.
9. Барер А.С., Сокол Е.А. Функциональное состояние зрительного анализатора человека – оператора. – М., 1990.
10. Зимкина Н.В. Физиология человека. – М., 1975.
11. Васильева В., Коссовская Э., Степочкина Н. Физиология человека. – М., 1973.
12. Физиология человека. / Под. Ред. Шмидта Р., Тевса Г. – Т1, М., 1996.
13. Общий курс физиологии человека и животных. – Т.1, М., 1991.
14. Основы физиологии человека. – Т.1, М., 1994.
| Физиология высшей нервной деятельности | |
|
Т.Г. Анищенко О.В. Глушковская-Семячкина Л.Н. Шорина Н.Б. Игошева ФИЗИОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Т.Г. Анищенко, О.В. Глушковская-Семячкина, Л ... Включение света воспринимается рецепторами сетчатки глаза собаки, и информация по зрительному нерву идет в головной мозг, достигая зрительного центра в коре больших полушарий ... Рецепторами зрительного анализатора являются палочки и колбочки, расположенные в сетчатке глаза, рецепторами слухового анализатора -чувствительные клетки кортиева органа ... |
Раздел: Рефераты по биологии Тип: реферат |
| Возрастная физиология и школьная гигиена | |
|
ВОПРОСЫ К ЗАЧЕТУ по возрастной физиологии и школьной гигиене Возрастная физиология 1. Предмет и задачи возрастной физиологии 2. Общие закономерности ... К первому типу экстерорецепторов относятся рецепторы, расположенные в коже (температурные и тактильные), и вкусовые, находящиеся в полости рта; ко второму - зрительные, слуховые и ... Например, зрительные рецепторы глаза возбуждаются при действии нескольких квантов света, рецепторы обоняния реагируют на действие двух-трех молекул пахучего вещества, а слуховые ... |
Раздел: Рефераты по медицине Тип: шпаргалка |
| Основные понятия анатомии и физиологии человека | |
|
1. Предмет и содержание анатомии и физиологии. Значение этих наук в психолого-педагогической подготовке будущего учителя Анатомия человека (от греч ... В зрительных буграх находятся нейроны, отростки которых идут к коре больших полушарий мозга. 1) рецепторов органа чувств, осуществляющих воспринимающую функцию (например, для зрительного анализатора это рецепторы сетчатки глаза); |
Раздел: Рефераты по биологии Тип: шпаргалка |
| Нервная ткань | |
|
Нервная ткань Нервная ткань (textus nervosus) - совокупность клеточных элементов, формирующих органы центральной и периферической нервной системы ... Чувствительные окончания принадлежат сенсорным нейронам, где рецепторный потенциал возникает под действием каких-либо раздражителей, внешних по отношению к нейрону стимулов. 3. Полисинаптические рефлекторные дуги с участием как спинного, так и головного мозга - в рефлекторных дугах этого типа имеется синапс в спинном мозге между сенсорным нейроном и ... |
Раздел: Рефераты по биологии Тип: курсовая работа |
| Особенности физической реабилитации у детей с нарушением зрения | |
|
ЗАБАЙКАЛЬСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ им. Н.Г.ЧЕРНЫШЕВСКОГО Факультет физической культуры Кафедра медико-биологических основ ... Затем, когда свет дойдет до фоторецепторов сетчатки и подействует на них, нервные импульсы, вызванные световым стимулом, должны пройти в обратном направлении через нервные волокна ... Эти аксоны, как уже было указано, являются немиелинизированными волокнами зрительного нерва, которые передают афферентные импульсы в зрительную кору большого мозга. |
Раздел: Рефераты по медицине Тип: дипломная работа |