Реферат: Факторы роста и выживание нейронов
Развивающиеся нейроны для своего выживания нуждаются в определенных белках, называемых факторами роста. Первым из факторов роста был идентифицирован фактор роста нерва (ФРН, NGF). В своих пионерских работах Леви-Монтальчини, Кохен с коллегами первыми показали, что ФРН стимулирует рост отростков сенсорных и симпатических нейронов. Позднее они показали, что ФРН необходим также для выживания нейронов. Блокирование действия ФРН у новорожденных мышат при помощи антител вело к гибели симпатических нейронов. При этом парасимпатическая система не поражалась, и спинномозговые ганглии были только немного меньше, чем в норме. В последующих экспериментах было показано, что на более ранней стадии развития ганглию также необходим ФРН для выживания и эти нейроны не выживали, если антитела вводились до рождения (путем иммунизации матери). У взрослых животных антитела к ФРН были гораздо менее эффективны в обоих популяциях клеток. Нуль-мутации ФРН и его высокоаффинного рецептора (что обсуждается ниже в этой главе) у трансгенных мышей также подтверждают, что симпатическим и сенсорным нейронам необходим ФРН для выживания. Таким образом, как симпатические, так и сенсорные нейроны имеют критический период своего развития, выживание в который зависит от наличия ФРН.
Захват и ретроградный транспорт ФРН
То, что от ФРН зависит выживание нейрона, говорит о том, что его действие происходит в теле клетки. Симпатические нейроны выращивают в специальных камерах, состоящих из трех отсеков (рис. 1). В центральном отсеке, куда помешаются нейроны, должен находиться ФРН, иначе нейроны гибнут. Однако, как только отростки нейрона достигают боковых отсеков камеры, ФРН может быть удален из центрального отсека и клетки остаются живыми, при условии, что в боковых отсеках ФРН остается (рис. 1В). Это дает основания полагать, что трофические эффекты ФРН на развивающиеся нейроны могут передаваться в виде ретроградного транспорта ФРН от нервных терминалей в тела клеток. Исследования, проведенные на взрослых животных с радиоактивно меченым ФРН, показали, что он активно захватывается в нервные терминали и активно транспортируется ретроградно в сому. Взрослым нейронам ФРН не нужен для выживания, однако он регулирует, среди всего прочего, синтез адреналина, индуцируя образование двух ферментов, необходимых для его синтеза: тирозингидроксилазу и дофамин гидроксилазу. При повреждении транспорта ФРН взрослого нейрона уровень этих ферментов падает.
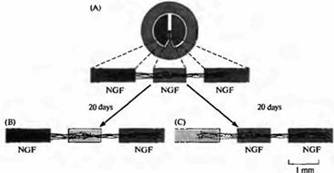
Рис. 1. Фактор роста нерва и выживание веточек аксонов клеток симпатических ганглиев, растущих в культуре клеток. (А) Нейроны, полученные из неонатального симпатического ганглия и помещенные в центральную секцию, посылают свои отростки под тефлоновым разделителем и в соседние секции; все секции содержат ФРН. (В) После начального роста отростков удаление ФРН из центральной секции в течение 20 дней не приводит ни к какому эффекту; нейроны в центральной секции используют ФРН, который ретроградно транспортируется из их терминалей, расположенных в боковых секциях. (С) После начального роста отростков удаление ФРН из левой секции вызывает дегенерацию нервных отростков в этой области,а те, которые находились в секции, содержащей ФРН, остаются интактными (Из Campenot, 1982.)
Факторы роста семейства нейротрофинов
Наблюдения, описанные в предыдущем разделе, привели к анализу молекулярных механизмов действия факторов роста большим количеством исследовательских групп, включая группы Леви-Монтальчини, Шутера, Тоенена и Барде). Они изучали такие вопросы, как свойства ФРН, рецепторов на мембране, которые связываются с ФРН, и последующие метаболические события. В слюнной железе ФРН присутствует в виде комплекса, состоящего из трех субъединиц. Ответственной за выживание нейрона и его рост является субъединица; она состоит их двух одинаковых белковых цепей, каждая из которых содержит 118 аминокислот и 3 дисульфидных мостика.
Сенсорные нейроны не только иннервируют свои мишени на периферии, но также направляют свои аксоны в ЦНС. Естественным является вопрос: существуют ли факторы, подобные ФРН, в ЦНС? В самом деле, в экстракте из ЦНС был обнаружен протеин, названный мозговым нейротрофическим фактором (МНФ, brain-derived neurotrophic factor, BDNF), который способствует выживанию нейронов дорзального спинного ганглия (ДСГ) в культуре и спасает их от гибели, если вводится in vivo эмбрионам во время периодов естественной гибели нейронов. Очистка и описание свойств МНФ выявили, что он имеет высокую степень гомологии с ФРН, показывая, что ФРН и МНФ являются членами одного семейства ростовых факторов, которое было названо нейротрофинами. Молекулярногенетические эксперименты помогли выявить еще несколько нейротрофинов: NT-3, NT-4/5 и NT-6. Все они состоят из димеров небольших основных белков, соединенных друг с другом при помощи бисульфидных связей между остатками цистеина. Димеры выглядят как симметричные белки с различными основными регионами, которые определяют свойства рецептора при встраивании его в мембрану (рис. 2).
На ранних периодах развития, до того, как произойдет иннервация периферических мишеней сенсорными нейронами, клеткам нервного гребня и сенсорным нейронам в ДСГ и ганглии тройничного нерва необходимы МНФ и NT-3 для пролиферации, дифференцировки и выживания. На таких ранних стадиях нейротрофины, по-видимому, секретируются самими нейронами, либо мезенхимальными тканями, расположенными вдоль роста аксонов сенсорных нейронов. В более поздние периоды, после того, как аксоны достигают своих целей, сенсорные нейроны начинают экспрессировать рецепторы к ФРН и становятся зависимыми от ФРН, секретируемого их мишенями.
Нейротрофины в ЦНС
Особенный интерес представляет нахождение популяции клеток, чувствительной к ФРН, в ЦНС. Таковыми являются холинергические нейроны, которые расположены в базальной области переднего мозга и которые иннервируют несколько различных структур, в том числе гиппокамп — область ЦНС, которая, как считают, связана с процессами памяти и обучения. Если аксоны этих нейронов у взрослой крысы перерезать, то клетки погибают. Однако, если после перерезания вводить в ЦНС ФРН, то клетки выживают после аксотомии. Число этих клеток, которые окрашиваются маркерами холинергических нейронов, уменьшается с возрастом, вместе со способностью крыс запоминать лабиринт и выполнять другие пространственные задачи. Если ФРН вводится взрослым крысам, число клеток, которые окрашиваются, увеличивается и улучшается выполнение крысами пространственных задач. Эти наблюдения показывают, что выживание и рост нейронов в ЦНС, скорее всего, зависит от факторов, которые одинаковы или близки к тем, которые были идентифицированы для периферических нейронов. Полученные результаты позволяют предполагать, в терминах молекулярной биологии, возможные дефекты этих факторов, которые могут привести к нарушениям развития мозга, и возможные пути борьбы с этими дефектами.
МНФ и NT-3 широко распространены в ЦНС как во время развития, так и у взрослых. Клетки коры и гиппокампа, скорее всего, нуждаются в МНФ и NT-3 для выживания, хотя изменения в выживаемости клеток в ЦНС трудно проследить. МНФ, как было показано, влияет на рост и сложность ветвления аксонов и дендритов в развивающейся ЦНС. Анализ мышей с генетическим дефицитом представляет многообещающий подход к пониманию роли нейротрофинов в развивающейся нервной системе .
Нейротрофины также могут играть определенную роль и в ЦНС взрослых животных. Физиологическая активность вызывает экспрессию определенных нейротрофинов в разных областях ЦНС и в ответ на различные стимулы. В свою очередь, нейротрофины, как было показано, способны влиять на физиологическую активность, облегчая синаптическую передачу и увеличивая или уменьшая возбудимость нейронов посредством регуляции экспрессии ионных каналов.
Рецепторы к нейротрофинам
Нейротрофины взаимодействуют с двумя типами рецепторов на поверхности нейронов--мишеней (см. рис. 3). Все нейротрофины связываются с относительно одинаковой и низкой активностью (Кd = 10--9 М) с мембранным рецептором, который был первоначально описан как низкоаффинный быстрый рецептор ФРН, или p75NGFR; более правильно называть этот рецептор низкоаффинным рецептором к нейротрофинам, или p75LNTR этот рецептор экспрессируется как в нейрональных, так и в ненейрональных клетках. Также существуют высокоаффинные (Кd = 10-11 М) рецепторы к нейротрофинам. Результаты биологических проб показывают, что эффекты нейротрофинов на выживание клеток и на рост отростков нервов обусловливаются высокоаффинными рецепторами.
Хотя высокоаффинные рецепторы к ФРН обычно расположены только на нейронах, они были первоначально обнаружены в клетках карциномы толстой кишки человека, как часть продукта синтеза онкогена trk (онкогенами называют гены, которые управляют трансформацией клеток). Аналог онкогена trk, находящийся в нормальных клетках, кодирует белок с массой около 140 кДа, называемый pl40prototrk или просто Trk. Структура белка Trk, предсказанная на основе его аминокислотной последовательности, состоит из внеклеточного домена, содержащего сайтдомен, кодирующий тирозинкиназу (рис. 3). Существуют по крайней мере три члена семейства про-онкогенов trk, каждый из которых является высокоаффинным рецептором для одного или нескольких нейротрофинов: TrkA — рецептор к ФРН и NT-6; TrkB, по видимому, является рецептором к МНФ и NT-4/5, a TrkC является рецептором к NT-3.
Связывание нейротрофинов с высокоактивными рецепторами приводит в первую очередь к увеличению фосфорилирования тирозина рецепторами. Это происходит вследствие аутофосфорилирования; вызванное лигандом образование рецепторного димера способствует приближению друг к другу внутриклеточных доменов тирозинкиназы каждого рецептора таким образом, что они фосфорилируют друг друга. Фосфорилирование рецептора Trk активирует четыре внутриклеточных сигнальных пути: фосфорилирование тирозина белка, названного SNT (suc-associated neurotrophic factor-induced tyrosine-phosphorylated target, связанная с suc и тирозин-фосфорилируемая в ответ на нейтрофические факторы мишень), фосфолипазу С, фосфоинозитол-3-киназу и каскад Ras-MAP киназы (рис. 4) .
Функция низкоаффинного рецептора, который не имеет внутриклеточных доменов, неизвестна. В некоторых клетках он может взаимодействовать с высокоаффинным рецептором во время связывания нейротрофинов. В других клетках, особенно в тех, в которых нет высокоаффинных рецепторов, он может управлять гибелью клеток или обеспечивать механизм ограниченной диффузии для установления высокой локальной концентрации нейтротрофинов, что необходимо для регенерации периферического нерва.
Конкурентные взаимодействия во время развития
После того как аксон достигает своих мишеней и образует синаптические связи, два процесса управляют паттерном иннервации. Одним является период гибели клеток, когда большое количество нейронов, которые установили синаптические связи, погибает. Вторым является уменьшение количества аксонов и синапсов, что сопровождается реорганизацией оставшихся связей, формирующих взрослый паттерн иннервации. Каждый из этих процессов регулируется конкуренцией за ограниченное количество факторов роста.
Гибель нейронов
Удивительной особенностью развития нервной системы является то, что многим ее нейронам суждено погибнуть во время развития. У беспозвоночных интенсивная гибель нейронов сопровождается стремительными изменениями, происходящими во время метаморфоза, и регулируется гормонами. Однако в развивающейся ЦНС как позвоночных, так и беспозвоночных гибель клеток происходит также и в отсутствие значительных морфологических изменений.
Эксперименты, выполненные Гамбургером и Леви-Монтальчини, впервые выявили запрограммированную гибель нейронов эмбрионов позвоночных, а также показали, что степень гибели клеток может зависеть от размеров ткани-мишени. Они показали, например, что в развивающейся конечности в то время, когда формируются первые синаптические связи с миофибриллами, от 40 до 70 % мотонейронов, которые послали аксоны в эту конечность, погибают. Имплантация дополнительной конечности уменьшает долю погибающих мотонейронов, а удаление одного из зачатков конечностей приводит к увеличению числа гибнущих клеток, что дает основания для предположения, что мотонейроны конкурируют за некие трофические вещества, выделяемые тканью-мишенью.
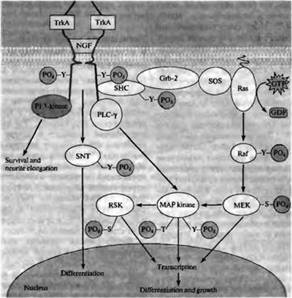
Рис. 3. Связывание димера фактора роста нервов с двумя TrkA рецепторами приводит к тому, что домен внутриклеточного белка тирозинкиназы каждого из TrkA рецепторов фосфорилирует остатки тирозина другого TrkA рецептора. Это запускает четыре внутриклеточных сигнальных каскада, которые приводят к росту и дифференцировке. Активация фосфоинозитол-3-киназы (PI 3-kinase) способствует удлинению отростков и их выживанию. Фосфорилирование тирозина SNT приводит к дифференцировке. Активация фосфолипазы C-γ (PLC-τ) стимулирует MAP киназы, которые индуцируют экспрессию генов, дифференцировку и рост как непосредственно, так и через фосфорилирование RSK. MAP киназа также активируется через сигнальный каскад, который включает связывание белков SHC, Grb-2, SOS и Ras, а также киназ Raf и МЕК. — S— Р04 — фосфорилирование серина; — Т — Р04 — фосфорилирование треонина; — Y — Р04 — фосфорилирование тирозина.
В самом деле, большое разнообразие нейротрофических белков, в основном вырабатываемых в мышце, как было обнаружено, могут управлять развитием мотонейронов: МНФ, NT-3 и NT-4/5, ИФР (инсулиноподобный фактор роста, IGF, insulin-like growth factor), CNTF (ciliary neurotrophic factor, нейротрофический фактор ресничек), GDNF (glial-derived neurotrophic factor, глиальный нейротрофический фактор), CDF (cholinergic differentiation factor, фактор холинергической дифференцировки, также называемый LIF, leukemia inhibitory factor, фактор подавляющий лейкемию) и ИФР-I (инсулиноподобный фактор роста 1). При введении их эмбриону эти белки предупреждают гибель мотонейронов, которые без этих факторов неминуемо погибли бы. Однако существует и несколько доказательств, включая анализ мутантных мышей, у которых отсутствует один или несколько подобных белков или их рецепторов, которые показывают, что ни один из этих факторов не является необходимым для выживания мотонейрона во время развития. Наилучшим кандидатом для фактора, регулирующего выживание мотонейрона, является еще пока неидентифицированный лиганд рецептора CNTF .
Избыточная продукция нейронов, после которой следует период гибели клеток, является характерной чертой развития нервной системы позвоночных. Некоторые из нейронов, которые погибают, возможно, не смогли образовать синапсов, или образовали их с неподходящей клеткой-мишенью. В подобных случаях гибель клеток связана с особенностью иннервации. Однако большинство клеток, которые гибнут, достигают и иннервируют правильные клетки-мишени. Таким образом, гибель клеток является основным механизмом, который поддерживает равновесие между количеством нейронов и их мишеней.
Неожиданной находкой явилось то, что ингибиторы образования мРНК или синтеза белка предотвращали гибель нейронов в отсутствие необходимых им нейротрофинов. Результаты этих и более поздних экспериментов показали, что гибель нейронов типично происходит путем апоптоза. Апоптоз — это процесс, который активирует внутреннюю «суицидальную» программу клетки, которая приводит к упорядоченному (запрограммированному) распаду ДНК и белков в клетке. Существенно, что для этого необходим предварительный синтез протеолитических ферментов или их активаторов.
Уменьшение числа связей и исчезновение полинейрональной иннервации
После того как популяция нейронов, иннервирующих определенную мишень, уменьшается благодаря гибели клеток, оставшиеся нейроны конкурируют друг с другом за синаптическую территорию. Это конкурирование обычно приводит к потере некоторых первично образованных веточек и синапсов («обрезка»). Уменьшение числа связей обеспечивает механизм для формирования правильной и полной иннервации мишени определенной популяцией нейронов. В некоторых случаях механизм уменьшения числа связей также обеспечивает механизмы коррекции ошибок; в других случаях он отражает стратегию поиска пути аксоном.
Особо яркий пример конкурентного уменьшения числа связей можно увидеть в развивающейся скелетной мышце. У взрослых животных каждый мотонейрон иннервирует группу до 300 мышечных волокон, формируя двигательную единицу , но каждое мышечное волокно иннервируется только одним аксоном. Однако в развивающейся мышце мотонейроны бурно ветвятся, так что каждое мышечное волокно в итоге иннервируется аксонами от нескольких мотонейронов (рис. 4), что называется полинейрональной иннервацией. В каждом развивающемся мышечном волокне синаптические окончания всех аксонов разбросаны вокруг одного участка, вблизи скоплений рецепторов АХ и других компонентов постсинаптического аппарата. По мере развития веточки аксонов исчезают, что приводит в конечном итоге к формированию взрослого паттерна. Этот процесс не вызывает гибель клетки (которая обычно происходит на более ранних стадиях развития), а только уменьшение количества мышечных волокон, иннервируемых каждым из мотонейронов.
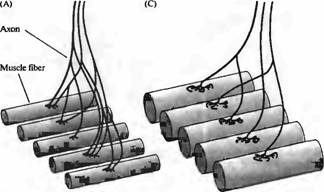
Рис. 4. (Рис. 4В, 4D см. на цветной вклейке после с. 640.) Полинейрональная иннервация и ее исчезновение в нерв но мышечном соединении позвоночных. (А) Во время развития эмбриона веточки аксонов моторных нейронов иннервируют множество мышечных волокон, и каждое мышечное волокно иннервируется несколькими мотонейронами (полинейрональная иннервация). (В) Флуоресцентная микрограмма нервно-мышечного соединения мыши (стадия Е18), показывающая распределение терминалей двух аксонов, каждый из которых помечен при помощи липофильной метки. В период полинейрональной иннервации терминальные ветвления аксонов мотонейронов, иннервирующих определенное мышечное волокно, переплетаются в областях отдельных синапсов. (С) После рождения полинейрональная иннервация исчезает по мере того, как веточки аксонов подвергаются ретракции, в результате чего каждое мышечное волокно получает иннервацию только от одного мотонейрона. (D) Флуоресцентная микрофотография нервно-мышечного соединения мыши во время исчезновения полинейрональной иннервации. Два аксона, иннервирующих синапс, были помечены как в В. Все терминали одного аксона исчезли, и сам аксон подвергся ретракции.
Исчезновение полинейронной иннервации регулируется конкуренцией между аксонами различных мотонейронов за синаптическое пространство на мышечных клетках. Наиболее ярким примером являются эксперименты на развивающихся мышцах лапки крысы. Когда все кроме одного моторные аксоны, иннервирующие эту мышцу, были перерезаны на раннем этапе развития, оставшийся аксон подвергся ветвлению и начал иннервировать большое количество волокон данной мышцы. В то время, когда обычно происходит исчезновение полинейрональной иннервации, ни один синапс не исчез. В отсутствие конкуренции для выживания мотонейрон поддерживает контакты с каждой миофибриллой, которую он ранее иннервировал. В экспериментах Лихтмана с коллегами были получены яркие картинки этого процесса при помощи визуализирования нервных терминалеи у животных in vivo при помоши витальных красителей и наблюдения за изменениями синаптической структуры во время исчезновения синапсов. Подобная же ретракция избыточного количества связей была показана в автономном ганглии неонатальных крыс и морских свинок. Каждая ганглионарная клетка первоначально получает большое количество связей, больше пяти, однако к возрасту 5 недель из них обычно остается только одна.
Активность нервов и исчезновение синапсов
Физиологические эксперименты показывают, что активность нейронов играет определенную роль в исчезновении синапсов, влияя как на интенсивность, так и на и результат конкуренции между терминалями аксонов. Стимулирование мышцы через нерв при помощи имплантированных металлических электродов увеличивает интенсивность исчезновения синапсов. Уменьшение активности путем добавления тетродотоксина в пространство вокруг нерва для блокирования потенциалов действия приводит к ингибированию синаптической передачи и замедлению исчезновения синапсов. На мышцах, которые получают сигналы от аксонов двух различных нервов, возможно проведение интересных экспериментов по блокированию проведения импульсов избирательно в одном из нервов. В таких случаях неактивные нейроны не могут, очевидно, нормально конкурировать: аксоны блокированных нейронов иннервируют двигательных единиц меньше, чем в норме; аксоны активных нервов иннервируют волокон больше, чем обычно. Однако доминирование неблокированных нервных волокон не полное, что говорит о роли других факторов, кроме активности, в исчезновении синапсов.
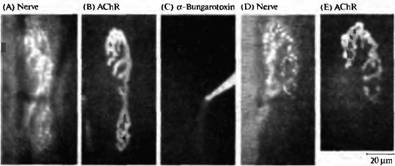
Рис. 5. Конкуренция между веточками одного мотонейрона, зависящая от активности. (А, В) Флуоресцентные микрофотографии нервно-мышечного соединения грудинососцевидной мышцы мыши, показывающие пресинаптическую терминаль (А), окрашенную при помощи 4-Di-2-Asp (который окрашивает митохондрии) и постсинаптическую мембрану (В), окрашенную при помощи родамин-связанного бунгаротоксина в низкой дозе (который окрашивает рецепторы АХ). (С) Рецепторы АХ в нижней части соединения блокированы при помощи насыщающей дозы немеченного -бунгаротоксина, что приводит к блокаде нервно-мышечной передачи селективно в этой области синапса. (D, Е) Через 31 день тот же самый синапс, что показан на А и В, был заново исследован. Терминаль аксона (D) и постсинаптические рецепторы (Е) исчезли из блокированной области. Таким образом локальная блокада передачи приводит к локальному исчезновению синапса.
Конкуренция в зависимости от активности также происходит на уровне веточек отдельного аксона двигательного нейрона. Если в небольшой области зрелого синапса произвести точечную аппликацию бунгаротоксина, то неактивный регион синапса подвергается элиминации (рис. 5). Если же блокировать весь синапс, его исчезновения не происходит. Молекулярные механизмы этой конкуренции, а также механизмы, посредством которых активность влияет на исчезновение синапсов, пока не открыты.
Сходная конкуренция В этом эксперименте среда во всех трех секциях изначально содержала ФРН. Клетки помещались в центральную секцию, а их отростки направлялись в каждую из двух боковых секций. ФРН затем удалялся из одной из боковых секций. Аксоны в центральной секции и в боковой секции, которая содержала ФРН, выживали, а аксоны в секции, из которой был удален ФРН, подвергались дегенерации (рис. 4С). Таким образом, в пределах ткани-мишени конкуренция между терминалями аксона за ограниченное количество нейротрофинов может приводить к изменениям синаптического ветвления аксонов, способствовать росту одних веточек и дегенерации других.за синаптические мишени возникает во время развития нервных путей в ЦНС. Примером может служить образование глазодоминантных колонок в зрительной коре, где аксоны из ядра ЛКТ, несущие информацию от разных глаз, первоначально значительно перекрываются в слое 4 коры, но затем формируются колонки правого и левого глаза. Таким образом, паттерн активности терминалей от обоих глаз играет решающую роль в определении результата этой конкуренции.
Нейротрофины и уменьшение количества связей
Уменьшение количества связей может возникать вследствие конкуренции за ограниченное количество трофических факторов, вырабатываемых клеткой-мишенью. Например, при развитии зрительной коры добавление избыточного количества МНФ блокирует образование колонок глазного доминирования, предотвращая потерю веточек аксонами ЛКТ. Эксперименты на симпатических нейронах, выращиваемых в трехсекционных камерах, могут также быть примером того, что обеспечение нейротрофинами может определять выживание или гибель индивидуальных веточек нейронов.
Общие размышления о нейронной специфичности
В последнее время наблюдается значительный прогресс в нашем понимании того, как нервная клетка находит свои мишени и устанавливает связи. Однако когда начинаешь рассуждать о несчетном количестве связей, которые должны сформироваться во время развития нервной системы, проблема нервной специфичности кажется просто пугающей. Даже банальная аналогия имеет очень сильный эффект. Представим, что мы не знаем ничего об устройстве и работе почтовой системы. Глава этой книги о нервной системе, без иллюстраций, отправляется из Триеста, Италия в Сандерланд, Массачусетс, куда она доходит за несколько дней. Как это происходит? Автор этой книги знает только ближайший к нему почтовый ящик и даже не знает, где находится почтовое отделение его района. Почтовый работник, который вынимает письма из ящика, знает, где находится почтовое отделение; там есть клерк, который может не знать, где находится Сандерланд, однако он знает, что посылку надо послать в аэропорт, и так далее, в нужную страну, город, улицу, здание и в конце концов конкретному человеку. Если этого не достаточно, то иллюстрации к главе высылаются из Денвера и Балтимора по тому же адресу, и они приходят почти одновременно с текстом из Триеста. После чего почта идет через те же самые почтовые ящики и отделение в обратном направлении к исходным отправителям.
Важной особенностью этой аналогии является то, что проблема кажется непостижимой на первый взгляд. Конечно, можно разгадать эту загадку с почтой, следуя за письмом шаг за шагом, пока оно не достигнет своего адресата. Это позволит выявить некоторые моменты в логике и структуре организации почты (не раскрывая сущность создателя этой системы). На каждом ее шаге выполняется ограниченное количество инструкций и используется ограниченное количество средств.
Некоторые механизмы нервной специфичности могут быть похожи на эту систему. Ганглиозная клетка сетчатки посылает свои отростки за пределы глаза, где они образуют зрительный нерв вместе с отростками других ганглиозных клеток сетчатки. Зрительный перекрест представляет следующую точку выбора, где решение о том, к какому, правому или левому, ЛКТ должен направляться отросток, может быть принято на основе химических сигналов. В ЛКТ аксоны сетчатки могут организовываться и иннервировать мишени основываясь на градиенте молекул-репеллентов. Аксоны нейронов ЛКТ подобным же образом следуют к своим мишеням в коре, останавливаясь на пути для формирования временных связей с промежуточными нейронами. Таким образом, задача по формированию специфических связей между ганглиозными клетками сетчатки и нейронами зрительной коры, кажущаяся такой сложной, может быть разбита на серию относительно простых, независимых шагов. Более того, при формировании связей во время развития расстояния между шагами очень малы; пути значительно удлиняются по мере созревания нервной системы.
Выводы
∙ В ЦНС позвоночных судьба развивающихся нейронов зависит в первую очередь от их расположения в росгрокаудальном направлении и уже затем от расположения в дорзовентральном направлении. Нох семейство гомеотических генов определяет идентифицирование клеток в зависимости от расположения в рострокаудальном направлении в заднем мозге. Белок, называемый Sonic hedgehog («Звучащий ежик»), синтезируемый в области хорды, управляет судьбой клеток, расположенных вентрально вдоль нервной трубки.
∙ Сигналы, управляющие дифференцировкой клеток, очень часто передаются через рецептор, связанный с тирозинкиназой, активирующей сложные внутриклеточные сигнальные каскады, которые приводят к изменениям в экспрессии генов. Примером подобной регуляции является развитие фоторецептора Drosophila.
∙ В коре ГМ млекопитающих развитие происходит таким способом, что нейроны, расположенные в более глубоких слоях, рождаются первыми.
∙ Нейроны нервного гребня возникают у края нервной складки и мигрируют из нервной трубки, образуя ПНС, пигментные клетки, а также кости черепа.
∙ Постоянное деление стволовых нервных клеток в ЦНС взрослых птиц и млекопитающих приводит к образованию новых нервных клеток.
∙ Кончик растущего аксона вытягивается, образуя конус роста.
∙ Поверхность клеток и молекулы адгезии, расположенные во внеклеточном матриксе, управляют конусом роста при помощи привлекающих и отталкивающих механизмов, действующих на короткой дистанции.
∙ Нетрины играют роль хемоаттрактантов, а семафорины — хеморепеллентов, действующих на большой дистанции, для многих типов аксонов.
∙ Эфрины (Ephrines) и Eph семейство рецепторов, связанных с тирозинкиназой, при помощи хемореппелентных механизмов управляют процессом поиска пути нервом, миграцией клеток и образованием связей между клетками.
∙ При соприкосновении конуса роста с мышечной клеткой в течение нескольких последующих минут устанавливается функционирующая нервно-мышечная передача.
∙ Выделение агрина из пресинаптических терминалей вызывает образование постсинаптической специализации в скелетной мышце.
∙ Нейронам для выживания и дифференцировки необходимы различные трофические факторы.
∙ Запрограммированная гибель нейронов довольно частое явление в развитии нервной системы.
∙ Синаптические связи, установленные однажды, в дальнейшем могут изменяться для обеспечения надежной и полной иннервации своей мишени. Перемоделирование связей осуществляется на основе конкуренции между терминалями аксона за нейротрофические факторы, выделяемые клеткой-мишенью в зависимости от активности терминалей.
Выводы
∙ В ЦНС позвоночных судьба развивающихся нейронов зависит в первую очередь от их расположения в росгрокаудальном направлении и уже затем от расположения в дорзовентральном направлении. Нох семейство гомеотических генов определяет идентифицирование клеток в зависимости от расположения в рострокаудальном направлении в заднем мозге. Белок, называемый Sonic hedgehog («Звучащий ежик»), синтезируемый в области хорды, управляет судьбой клеток, расположенных вентрально вдоль нервной трубки.
∙ Сигналы, управляющие дифференцировкой клеток, очень часто передаются через рецептор, связанный с тирозинкиназой, активирующей сложные внутриклеточные сигнальные каскады, которые приводят к изменениям в экспрессии генов. Примером подобной регуляции является развитие фоторецептора Drosophila.
∙ В коре ГМ млекопитающих развитие происходит таким способом, что нейроны, расположенные в более глубоких слоях, рождаются первыми.
∙ Нейроны нервного гребня возникают у края нервной складки и мигрируют из нервной трубки, образуя ПНС, пигментные клетки, а также кости черепа.
∙ Постоянное деление стволовых нервных клеток в ЦНС взрослых птиц и млекопитающих приводит к образованию новых нервных клеток.
∙ Кончик растущего аксона вытягивается, образуя конус роста.
∙ Поверхность клеток и молекулы адгезии, расположенные во внеклеточном матриксе, управляют конусом роста при помощи привлекающих и отталкивающих механизмов, действующих на короткой дистанции.
∙ Нетрины играют роль хемоаттрактантов, а семафорины — хеморепеллентов, действующих на большой дистанции, для многих типов аксонов.
∙ Эфрины (Ephrines) и Eph семейство рецепторов, связанных с тирозинкиназой, при помощи хемореппелентных механизмов управляют процессом поиска пути нервом, миграцией клеток и образованием связей между клетками.
∙ При соприкосновении конуса роста с мышечной клеткой в течение нескольких последующих минут устанавливается функционирующая нервно-мышечная передача.
∙ Выделение агрина из пресинаптических терминалей вызывает образование постсинаптической специализации в скелетной мышце.
∙ Нейронам для выживания и дифференцировки необходимы различные трофические факторы.
∙ Запрограммированная гибель нейронов довольно частое явление в развитии нервной системы.
∙ Синаптические связи, установленные однажды, в дальнейшем могут изменяться для обеспечения надежной и полной иннервации своей мишени. Перемоделирование связей осуществляется на основе конкуренции между терминалями аксона за нейротрофические факторы, выделяемые клеткой-мишенью в зависимости от активности терминалей.