Курсовая работа: Життєві форми організмів
Зміст
Вступ. 2
Розділ 1. Життєві форми та підходи до їх класифікації 3
1.1 Поняття про біоморфу. 3
1.2 Основні підходи до класифікації життєвих форм. 4
Розділ 2. Характеристика життєвих форм за Серебряковим. 14
2.1 Дерева. 14
2.2 Чагарники. 17
2.3 Чагарники. 19
2.4 Напівчагарники і напівчагарники. 21
Висновок. 30
Список використаної літератури. 31
Вступ
Життєва форма визначається як пристосованість організмів до комплексу факторів середовища та певного способу життя. Вперше життєві форми були виділені у рослин. Ще Теофраст класифікував рослини, виділяючи серед них дерева, чагарники, напівчагарники, трав'янисті рослини. Термін "життєва форма" введений в 1884 р. датським ботаніком Е. Вармінгом, під яким він розумів "форму, в якій вегетативне тіло рослини знаходиться в гармонії з зовнішнім середовищем протягом всього життя, від насіння до відмирання." Як синоніми цьому терміну в екології рослин використовуються "екобіоморфа", "біотип", "форма росту", "епіморфа".
Вивченням життєвих форм організмів займалися багато вчених ботаніків. Існує кілька класифікацій життєвих форм, в основу яких покладено різні ознаки, як анатомічні так і фізіологічні.
Предмет дослідження – підходи до класифікацій різних авторів.
Об`єкт дослідження – життєві форми організмів.
Мета нашого дослідження – вивчити різновиди життєвих форм на дію різних екологічних факторів.
Завдання роботи:
дати поняття про життєву форму (біоморфу);
розглянути бачення різних вчених-ботаніків класифікації життєвих форм організмів;
розглянути класифікацію життєвих форм за Серебряковим.
Розділ 1. Життєві форми та підходи до їх класифікації
1.1 Поняття про біоморфу
Біоморфа — життєва форма, що визначається систематичним становищем видів, їх формами росту і біологічними ритмами (циклічними коливаннями біологічних процесів і явищ). Біоморфи визначають пристосованість організмів до комплексу факторів і зумовлені специфічними умовами їх місцезростання. Характеризуються вони габітусом — своєрідним зовнішнім виглядом, що сформувався в процесі онтогенезу під впливом умов середовища і насамперед ґрунтово-кліматичних.
Морфологічні зміни пов'язані з функціонуванням організму і його органів, що дає можливість особинам оптимально використовувати зовнішнє середовище і пристосовуватись до нього. Отже, функціональність форми та її адаптивний характер — це дві сторони одного й того самого явища.
Перша класифікація рослин за біоморфами була запропонована ще О. Гумбольдтом в праці "Ідеї географії рослин", де він виділяє 17 основних форм рослин: банани, пальми, деревовидні папороті, орхідеї, мімозові тощо. Вчений виділяв форми на основі фізіономічних ознак, не заглиблюючись в екологічні особливості формування життєвих форм, а тому сюди потрапили й деякі групи нижчих рослин — гриби й лишайники.
Згодом форми і функції рослин були покладені в ряд наступних класифікацій рослинності. Розроблялися численні набори символів для опису таких ознак, як розміри рослини, життєва форма, морфологія, розміри і текстура листя, відсоток покриття. Наприклад, метод, запропонований П'єром Дансеро, описує рослини за допомогою різноманітних фігур.
1.2 Основні підходи до класифікації життєвих форм
У 1903 р. датський ботанік Крістен Раункієр запропонував систему класифікації рослин, засновану на розташуванні їх бруньок поновлення, і виявив, що поширення головних виділених ним категорій добре співпадає з розподілом кліматичних умов. Раункієр виділив п'ять основних життєвих форм (рис. 1). Класифікація життєвих форм, за Раункієром, така:
1. Фанерофіти (від грецького phaneros — явний) — бруньки поновлення розміщені високо над поверхнею землі (дерева, кущі). Бруньки захищені лусками.
2. Хамефіти — бруньки поновлення розміщені близько від поверхні землі. Бруньки захищені не тільки лусками, а й снігом та відмерлим листям (журавлина, брусниця, карликова береза).
3. Гемікриптофіти — бруньки поновлення розміщені на рівні поверхні землі. Тому бруньки захищені не тільки лусками, а й снігом, відмерлим листям та грунтом (кульбаба, підбіл).
4. Криптофіти — бруньки поновлення розміщені під поверхнею землі (рослини, що мають цибулини, бульби, кореневища).
5. Терофіти — однорічники, що перезимовують у вигляді насіння. Раункієр вважав, що життєві форми складаються історично як результат пристосування рослин до кліматичних умов середовища. Відсоткове співвідношення і розподіл видів за життєвими формами в даному угрупованні в регіоні досліджень він назвав біологічним спектром. Для різних кліматичних зон були складені біологічні спектри, які були індикаторами клімату. Так, спекотний і вологий клімат тропіків був названий кліматом фанерофітів, континентальний клімат помірної зони - кліматом гемікриптофітів.
Порівнюючи рослини різних районів. Раункієр вивів таку закономірність: холодні пояси багаті на хамефіти; помірний пояс багатий на гемікриптофіти; у тропічних лісах переважають фанерофіти. Недоліком системи Раункієра є недостатній історичний зміст її.
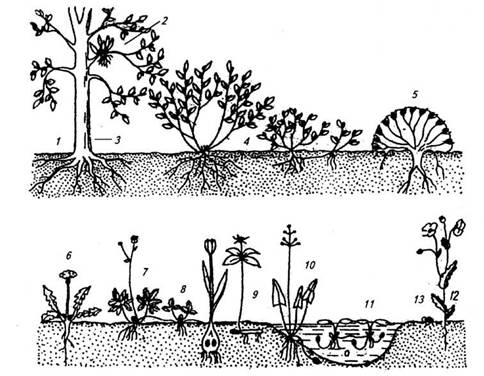
Рис. 1. Життєві форми рослин за Раункієром: 1-3 — фанерофіти; 4, 5 — хамефіти; 6, 7 — гемікриптофіти; 8-11 — криптофіти; 12 — терофіти; 13 — насіння із зародком.
Система життєвих форм, розроблена О.І. Зозулиним, має історичний і генетичний зміст. За визначенням О.І. Зозуліна. життєві форми складаються з сукупності рослинних організмів, які мають якісно подібні пристосування для вдержання ними площі населеності і розростання на цій території. Отже, вона витримана у фітоценологічному плані. Всі рослини згідно системи Зозулина розбиває на чотири життєві форми:
1. Редитивні рослини (від лат. reddere — поступатися) — багаторічні види, які поступаються під час росту особин своїм місцем у випадку знищення з якої-небудь причини надземної частини (багато дерев тропічної зони, в яких немає сплячих бруньок для основи стовбура. У наших умовах до таких можна віднести більшість хвойних порід).
2. Рестативні рослини (від лат. restore — залишитися) — багаторічні види, які поновлюються від сплячих бруньок у разі знищення надземних частин (це більшість дерев, кущів і трав помірного поясу).
3. Іруптивні рослини (від лат. ітітрге — вторгатися) — багаторічні види, які вторгаються за допомогою нових пагонів у місця, зайняті іншими видами (це переважно кореневищні і коренепаросткові рослини).
4. Вагативні рослини (від лат. vagare — кочувати) — однорічні і дворічні види, які розмножуються лише насінням (не вдержують за собою площу), за допомогою вітру, який його розносить. На новому місці насіння проростає.
Ці типи діляться ще на постійно вічнозелені, постійно листопадні і періодичні. Єдність принципу класифікації життєвих форм надає системі Зозулина риси цілеспрямованості і системності. Але це штучна система тому, що дерева віднесені до трьох типів. До другого типу віднесено дерева, кущі і трави.
Російським ботаніком І.С.Серебряковим була запропонована класифікація життєвих форм на основі архітектоніки вегетативних органів. Життєва форма за Серебряковим, це своєрідний зовнішній вигляд (габітус) певної групи рослин, який формується в процесі онтогенезу, росту і розвитку в певних умовах середовища. Життєві форми складаються під впливом сприятливих і несприятливих умов середовища. Серебряков використав велику сукупність ознак в системі співвіднесення, виділивши такі одиниці класифікації: відділи, типи, класи, підкласи, групи, підгрупи, секції життєвих форм та власне життєві форми.
І.С. Серебряков виділяє відділи: А — наземні і епіфітні рослини, які включають три типи: 1) дерева, 2) кущі, 3) кущики.
Відділ В — напівдеревні рослини (IV тип — напівкущі і напівкущики).
Відділ С — наземні трави (V—тип полікарпічні трави, VI тип— монокарпічні трави).
Відділ D — водяні трави (VII тип — земноводні трави, VIII тип — плаваючі і підводні трави).
Таким чином, Серебряков поділяє відділи насамперед за структурою надземних осей (деревні, напівдеревні і трав'янисті). У відділі деревних рослинвін виділяє типи за відносною тривалістю життя надземних осей; у відділі наземних трав'янистих рослин — за тривалістю життя рослин у цілому.
Послідовність відділів і типів у системі Серебрякова показує основні напрями еволюції життєвих форм покритонасінних від дерев до трав. Класи в межах типу виділяються за ознакою більшого екологічного і еволюційного значення, що виявлено в структурі пагонів (рослини з повзучими, розетковими та іншими стеблами). У ряді типів виділяються класи на основі спеціалізації живлення рослин (сапрофіти і паразити), способу життя (епіфіти) тощо. Але це не порушує принципу, бо своєрідність живлення способу життя цих рослин знаходить свій вияв у структурі їх пагонів. Особливості структури пагонів, за Серебряковим, надають життєвим формам кожного класу різко виявленої морфологічної і економічної своєрідності. Закінчується система життєвих форм типом монокарпічних трав'янистих рослин.
Біоморфологічна структура визначається як властиве флорі кількісне співвідношення елементів, які різняться за біоморфологічними ознаками. Як було показано ще О.Гумбольдтом, Х.Стевеном та К.Раункієром, таке співвідношення для подібних флор характеризується високою константністю та являє їх важливу характеристику. Як відмічають Г.М. Зозулін, І.Г.Серебряков, В.М.Голубєв. С.М. Зиман, сучасні системи життєвих форм є еволюційними та суттєво різняться між собою у зв'язку з різними принципами, покладеними у їх основу.
Біоморфологічна структура є важливим показником регіональної флори, оскільки в значній мірі залежить від екологічних умов, насамперед від грунтово-кліматичних та екотопічних. Складні системи взаємовідносин рослин та умов оточуючого середовища виявляються у фізіологічних особливостях і життєвій формі (габітусі) рослин.
Вивчення життєвих форм різних фітоценозів сприяє більш глибокому пізнанню структури, динаміки рослинних співтовариств і їхнього взаємозв'язку із середовищем. По Гамсу, життєві форми є адекватним вираженням умов життя організмів; тому їхнє вивчення в біоценозах - надійний засіб екологічної оцінки місце існування.
Б. А. Келлер підкреслював, що розчленовування рослинного покриву на життєві форми і з'ясування їхньої ролі дає більш глибоке представлення про будівлю рослинних угруповань і про їхні відносини із середовищем.
По Адамсу, специфіка біологічного спектра тієї або іншої країни визначається її історією. Дослідження біологічних спектрів різних пустель Азії, Африки, Америки й Австралії приводять цього автора до висновку, що при подібності клімату часто виявляється значне розходження в складі життєвих форм, що порозумівається особливостями флори і її походженням.
Кейн йде далі і дійде висновку, що спектри окремих асоціацій тонко реагують на особливості мікроклімату. Тонка реакція спектра життєвих форм біоценозів на зміну умов виростання може бути використана при вивченні екологічних рядів асоціацій.
Е. М Лавренко показав, що різноманітний склад життєвих форм у рослинному співтоваристві забезпечує усебічне використання живими істотами життєвих ресурсів місця існування, більш-менш стійкий максимум рослинної продукції і найбільший ступінь впливу на середовище, тобто її біогенне перетворення. У цьому зв'язку вивчення життєвих форм здобуває велике значення. При вивченні життєвих форм рослин велика увага дослідників займала і займає проблема їхньої класифікації.
Ряд систем було запропоновано радянськими, російськими й українськими ботаніками ботаніками. В. Н. Сукачов для ялинових лісів виділяє характерні групи життєвих форм:
1) тіньовитривалі мікотрофні хвойні,
2) трав'янисті довгокореневищні мікотрофні рослини трав'яного типу,
3) довгокореневищні трав'янисті мікотрофні з листами брусничного типу,
4) злаковидні зимньо-зелені рослини тіньового типу,
5) тіньові трави з клейстогамними квітками,
6) вічнозелені голколисті мікотрофні трави,
7) зимньо-зелені мікотрофні чагарнички,
8) літньо-зелені чагарнички,
9) напівсапрофіти,
10) сапрофіти,
11) напівпаразити.
Виділення цих груп засновано на принципах з урахуванням способу живлення, сінності розвитку листя, наявності або відсутності кореневищ, ознак фізіологічного характеру. Відзначені Сукачовим групи відбивають різноманіття форм і шляхів пристосування рослин до умов виростання в тінистих ялинових лісах.
Також велика увага приділяється класифікаціям життєвих форм пустель, напівпустель, степової зони. Б.А. Келлер виділив у степах, пустелях, напівпустелях:
1) багаторічні двочасткові трави,
2) багаторічні кущові і корневищні трави,
3) багаторічні дереново-степові злаки з вузькими листками,
4) карликові напівчагарнички,
5) весняні ефемери,
6) нижчі рослини на ґрунті.
Більш детальна система життєвих форм для степової рослинності розроблена В.В. Алехиним (1936):
А. Чагарники і напівчагарники.
1. Степові чагарники.
2. Напівчагарники.
Б. Трав'янисті дворічники і багатолітники.
3. Рослини з листами по всій довжині стебла; стебло вертикальне, міцне
1) Високі рослини верхнього степового ярусу.
2) Рослини середнього і більш низького ярусів.
4. Рослини з лисктами, зближеними в поверхні землі в розетки.
5. Рослини, що утворять перекотиполе.
6. Рослини цибулинні і бульбові ефемери.
7. Рослини цибулинні літнього циклу.
8. Дерновинні злаковидні рослини з вузькими листками (злаки й осоки).
9. Недерновинні злаковидні рослини з більш широкими листами.
В. Однорічники.
10. Весняні ефемери.
11. Літньо-осінні однолітники.
12. Мохи.
13. Лишайники.
14. Водорості.
Система В.В.Алехина залишається однієї з найбільш розроблених для наших степів. В.С. Закржевским і Е.П.Коровиним дається перелік екологічних форм рослин для глинистих пустель Бет-Пакдала:
1. Група склерофільних чагарників.
2. Група ксерофільних напівчагарників.
1) Форма афільна або зі скороченими листами.
2) Форма з ефемерними листами.
3) Форма із сукулентними листками.
4) Форма зі склероморфними листками.
5) Форма з плоскими листами.
3. Група трав'янистих сукулентних форм.
4. Група ефемерних трав.
1) Ефемери.
2) Ефемероїди.
В.С.Закржевский і Е.П.Коровин відзначають, що кожна з названих форм володіє своїми біологічними особливостями. Вони знайшли визначену екологічну приуроченість виділених груп рослин до різних умовах у межах Бет-Пак-Далаэ
Для Каракумів М.П.Петров пропонує наступну систему:
А. Фанерофіти.
Б. Хамефіти.
В. Гемікриптофіти.
1) Період спокою не залежить від недоліку води.
2) Період спокою залежить від недоліку води.
М.Криптофіти.
1) Вегетативні органи завжди у воді.
2) Вегетативні органи земноводних.
3) Вегетативні органи в землі - гелофіти: автотрофні (1-період спокою не залежить від недоліку води, 2-період спокою залежить від недоліку води) і гетеротрофні.
М.В.Сенянинова-Корчагина (1949) при аналізі фітоклімату Ленінградської області бере за основу систему життєвих форм Раункієра, доповнюючи неї. Нею виділяються 20 життєвих форм:
1) фанерофіти вічнозелені,
2) фанерофіти літньозелені,
3) хамефіти вічнозелені,
4) хамефіти літньозелені,
5) гемікриптофіти вічнозелені розеточні з кореневищем,
6) гемікриптофіти вічнозелені розеточніз коренем,
7) гемікриптофіти вічнозелені з пагонами,
8) гемікриптофіти вічнозелені дерновидні,
9) гемікриптофіти вічнозелені рихлокущові,
10) гемікриптофіти літньозелені розеткові і рихлокущові,
11) гемікриптофіти літньозелені розеточні і дернинні з коренями,
12) гемікриптофіти літньозелені з пагонами,
13) гемікриптофіти літньо зелені, що зимують тільки бруньками на кореневище,
14) гемікриптофіти літньозелені, що зимують тільки бруньками на корені,
15) геофіти корневищні,
16) геофіти цибулинно-корневищні,
17) геофіти з бульбами,
18) гелофіти або гігрофіти з кореневищами,
19) гідрофіти,
20) терофіти.
Автор говорить, що серед гемікриптофітів є ряд добре помітних морфологічних і екологічних форм, характерних для різних асоціацій рослинності з особливим екологічним режимом.
Детальна система життєвих форм степових рослин розроблена М.С.Шалітом (1955). В основу цієї системи покладено кілька ознак: форма росту, тривалість життя, кількість плодоносінь, спосіб вегетативного розмноження, характер кореневої системи. Квіткові рослини в ній розділяються на багатолітники і однорічники. Серед полікарликів виділені:
1) чагарники і чагарнички,
2) напівчагарники і напівчагарнички,
3) напівтрави,
4) трави, триваловегетуючі і коротковегетуючі.
При подальшому підрозділі виділяються більш вузькі групи осьових рослин і повзучих рослин, дернистих рослин, групи сукулентів, перекотиполе, подушковидних форм.
У перерахованих системах життєвих форм за основу прийняті різні екологічні, фізіологічні, і біологічні ознаки рослин: форма росту, тривалість життя, ритм сезонного, розвитку, характер харчування; здатність рослин до вегетативного поновлення і розмноження. Ці ознаки мають важливе фітоценотичне значення. У багаторічних рослин з інтенсивністю вегетативного поновлення і розмноження часто зв'язаний ступінь участі виду в співтоваристві.
Розділ 2. Характеристика життєвих форм за Серебряковим
2.1 Дерева
1-й клас. Кроноутворюючі дерева з цілком здеревілими подовженими пагонами.
Стовбур сильно гілчастий, що несе у верхній частині крону з гілок різного порядку (вегетативних і квіткових).
Дерева цього підкласу в плині всього життя зв'язані з ґрунтом кореневою системою.
Група А. Дерева з "звичайною", лише підземною кореневою системою. Серед них виділяються три підгрупи.
Підгрупа а. Дерева з прямостоящими стовбурами. Ріст стовбурів завжди ортотропний. До них відносяться п'ять секцій.
Секція 1. Одностовбурні дерева з високим стовбуром (лісового типу). Сюди відносяться, наприклад, Quercus robur, Betula pendula Roth, Tіlіa cordata Mіll. Дуже багато дерев вологих помірно-холодних областей, помірно-теплих і тропічних областей. По ритму розвитку листя дерева цієї секції підрозділяються на підсекції листопадних і лісових вічнозелених дерев з різною тривалістю життя листів.
Секція 2. Кущовидні, або небагатостовбурні (плейокормні) дерева (субальпійського і субарктичного типу). Являють собою перехідні форми від типових дерев до чагарників. Від чагарників відрізняються великим довголіттям кістякових осей і їх меншою кількістю. Найбільш характерні для субальпійської і субарктичної границі лісової рослинності. Гарним прикладом може служити Betula tortuosa.
Секція 3. Одностовбурні дерева з низьким стовбуром (дерева лісостепового і саванново-лісового типу) або "дерева плодового типу" (Schіmper u. Faber. 1935). В онтогенезі й архітектурі дерева є риси чагарників. До цієї секції відносяться: багато дерев лісів Африки, Південної Америки й Азії з різних сімейств двочасткових, а також маслини. Дерева цієї секції підрозділяються на підсекції листопадних і вічнозелених дерев з різною тривалістю життя листів.
Секція 4. Саванові дерева з водозапасаючим, різко потовщеним (іноді бочковидно), могутнім стовбуром. До цієї секції відносяться баобаб і каванілезія.
Секція 5. Дерева із сезонно сукулентними однолітніми безлистими пагонами. Це мешканці пустель і напівпустель з печенею влітку і морозною зимою.
Підгрупа б. Дерева з лежачим стовбуром, що укореняється, і головними гілками - стланці. Типовим прикладом таких дерев є Pіnus pumіla (Pall.) Rgl Junіperus turkestanіca, у субальпійському поясі деяких хребтів Тянь-Шаню.
Підгрупа в. Дерева з ліановидним стовбуром. Поширені винятково у вологих тропічних лісах. Сюди відносяться, наприклад, Lonchocarpus cyanescens Benth.
Група Б. Дерева володіють, крім "звичайних" підземних коренів, ще і спеціалізованими надземними коренями.
Підгрупа а. Дерева з "ходульними" надземними коренями в нижній частині стовбура. Це переважно дерева мангрових заростей і деякі невисокі дерева волого-тропічних лісів.
Підгрупа б. Дерева з "дихальними" коренями. Ці корені мають негативний геотропізм і висуваються з ґрунту. Зустрічаються в мангрових заростях, наприклад види Avіcennіa.
Підгрупа в. Дерева з "досковидними" виростами - підпірками кореневого походження в підставі стовбура. Це головним чином гігантські дерева влажнотропических лісів, наприклад Ceіba pentandra Gaeerthn.
2-й підклас. Геміепіфітні кроноутворюючі дерева.
Починають розвиток як типові епіфіти, але з часом повітряні корені їх досягають поверхні ґрунту, проникають у неї, сильно товщають над землею й утворять помилкові стовбури - "кореневі стовбури".
Група А. Вегетативно-нерухомі геміепіфітні кроноутворюючі дерева. Повітряні корені - помилкові стовбури ростуть лише вниз по стовбурі хазяїна. Сюди відносяться "деревья-душителі" різних типів, наприклад види Fіcus волого-тропічних лісів Південно-східної Азії.
Група Б. Вегетативно-рухливі геміепіфні кроноутворюючі дерева. Повітряні корені ростуть не тільки вниз по головному стовбурі, але й уздовж галузей крони дерева-хазяїна, утворити безліч колоновидних "стовбурів" кореневого походження. Сюди відносяться деякі види Fіcus волого-тропічних лісів Південно-східної Азії, наприклад банианы - "баніан форма" Грізебаха.
2-й клас. Розеточні дерева.
Дерева зі стовбуром слабко або зовсім негілчастим, що постійно утворить на своїй вершині лише укорочені втечі, що несуть розетки великих листів. На відміну від дерев попереднього класу, що утворять крону з гілок, розеточні дерева несуть листову крону. Недорозвинення міжвузля у розеточних дерев зв'язано зі слабким одеревенінням і недостатньою механічною міцністю стебла. Іноді, очевидно, це еволюційно древні форми, що збереглися в тропічних і субтропічних лісах.
1-й підклас. Негілчасті розеткові дерева
Крайні форми цього класу мають лише простий не гілчастий стовбур з розеткою листів на верхівці. Сюди відносяться види Ravenala, Strelіtzіa, некотрые Lobelіaceceae і ін.
2-й підклас. Гілчасті розеткові дерева
Утворять ряд форм від дуже слабко гілчасті, наприклад Xanthorrhaea preіssіі Endl., види Pandanus, до регулярно гілчастихся, наприклад, різні деревні лілейні Dracaena. Yucca. Cordylіne, деякі Composіtae і Aloe. 3-й підклас.
Сукулентно-стеблові безлисті дерева.
Дерева зі стовбуром і галузями (де вони є) м'ясистої консистенції, з укороченими міждвузлями і недорозвиненими листами, що нерідко перетворюється в колючки. Стебла виконують водозапасаючу й асимілюючу функцію. Мешканці тропічних і субтропічних аридних областей без холодного, (зимового) часу року. Сюди відносяться численні види Cactaceae. Euphorbіaceae і ряд інших.
2.2 Чагарники
Головний стовбур мається лише на початку життя рослини. З віком головний стовбур повністю відмирає. В дорослому стані чагарник володіє декількома або багатьма надземними кістяковими осями. Тривалість життя надземних кістякових осей у чагарників невелика: 10-12 років, хоча може бути від 2-3 років до 30-40 років і більш. Висота чагарників від 0,8-1,0 до 5-6 м. Широко поширені від екваторіальних областей до холодних зон. Чагарники розділяються на чотири класи.
1-й клас. Чагарники з цілком здеревілими подовженими погонами.
Цей клас аналогічний подібному до класу дерев. Сюди відноситься більшість чагарників, у яких кістякові надземні осі ортотропні (1-й підклас ), плагіотропні (2-й підклас) або ліановидні (3-й підклас), більш-менш рясно гілчасті, з подовженими міжвузлями річних пагонів.
1-й підклас. Прямостоячі чагарники.
Надземні осі ортотропні; вони утворять те більш-менш щільний пучок при слабкому розвитку їх плагіотропних підземних частин (як у ліщини) , те розташовуються рідко. Цей підклас чагарників найбільш багатий представниками, що поширені в різних як гумідних, так і аридних кліматичних областях.
Група А. Аероксильні чагарники. Розгалуження осей надземне. Перехідні форми між деревами і чагарниками.
Підгрупа а. Пухкі аероксильні чагарники. Прикладом можуть служити Sorbus aucuparіa, Padus racemosa у несприятливих заболочених місцеперебуваннях або густих лісах тайгової зони європейської частини Росії. У більш сприятливих умовах ці ж види утворять кущовидні дерева.
Підгрупа б. Подушковидні аероксильні чагарники. Характеризуються невеликих і однаковим для усіх утеч річним приростом і щільним розташуванням кістякових осей і галузей. До них відносяться: Bupleurum frutіcosum, Poterіum spіnosum, Astragalus.
Група Б. Геоксильні або дійсні чагарники. Рослини з підземним галуженням осей и утворенням довговічних підземних одеревенілих осей - ксилоподіїв, від яких відходять більш тонкі і менш довговічні надземні осі. Сюди відноситься більшість чагарників європейської флори. До цієї ж групи належать багато видів бамбуків Тропічних і субтропічних країн.
Аероксильні і геоксильні чагарники у свою чергу можуть бути підрозділені на секції вегетативнонерухомих і вегетативнорухомих за допомогою кореневих відростків і столонів.
2-й підклас . Напівпростратні і сланкі чагарники.
Головна вісь і бічні галуження лежачі, укорінені, піднімаються до верхівки. Більшість цих чагарників, ймовірно галузяться тільки надземно. Рідко зустрічаються в аридних областях, але виявляються домінуючими в субальпійській і субарктичній областях, утворити криволісся ( заросли верболозів у горах Скандинавії.
3-й підклас. Ліановидні чагарники.
Мають кістякові осі, нездатними до самостійного ортотропного росту і використовують сусідні деревні рослини як опору. Відрізняються від ліановидних дерев більш тонкими стеблами, від 1-2 до 5-8 см у діаметрі. Переважно мешканці вологих тропічних і субтропічних лісів. До цього підкласу відноситься більшість деревних ліан. Класифікація в межах підкласу ґрунтується на способах лазання.
2-й клас. Розеткові чагарники з укороченими пагонами.
Аналогічні класу розеткові дерев. Наземні осі з укороченими міжвузлями і розетками великих листів. Зустрічаються рідко. Сюди відносяться: Batrіcs major, Senecіostewartіae і ін.
3-й клас. Сукулентно-стеблові безлисті чагарники.
Кістякові осі м'ясистої консистенції виконують водозапасаючу й асиміляційну функції. Переважають прямостоячі форми (прикладом є кактуси).
4-й клас. Паразитарні і напівпаразитарні чагарники.
З ґрунтом не зв'язані. Живуть на інших деревних рослинах, присмоктуючи до них спеціальними органами - гаусторіями. У більшості випадків мають зелені листи і самі здатні до фотосинтезу. Сюди відносяться види Loranthaceae, Vіscum album, Osyrіs alba і ін.
2.3 Чагарники
Деревні рослини, у яких головна вісь мається лише на початку онтогенезу, а потім відмирає і змінюється бічними надземними осями зі сплячих бруньок базальної частини материнської осі. Укорінені бічні кістякові осі гілкуються, утворити парціальні кущі. Тривалість життєвого циклу ортотропних надземних осей у чагарничків не перевищує 5-10 років, а висота рослин коливається від 5-7 до 50-60 див. Переважно мешканці високогірних областей, помірно-холодних і холодних зон. Підрозділяються на 4 класи.
1-й клас. Чагарнички з повністю одеревенілими подовженими пагонами.
Надземні осі більш-менш рясно галузяться. Річні пагони мають подовжені міжвузля.
1-й підклас. Вегетативно-нерухомі, прямостоячі чагарнички з цілком здеревілими подовженими пагонами.
Утворять форми з тісно зближеними надземними осями, без довгих підземних ризомів; нездатні до вегетативного розмноження.
Група А. Аероксильні вегетонерухомі чагарнички. Розгалуження мається лише в надземній частині, біля поверхні ґрунту.
Підгрупа а. Пухкі аероксильні чагарнички. По Дюрье такі чагарнички досить звичайні серед заростей чагарничків Південної Австралії., у Сіднею, на острові Хуан Фернандес.
Підгрупа 6. Подушковидні аероксильні чагарнички. Річний приріст усіх пагонів однаковий і незначний по величині. Поширені в субальпійських і альпійських, субантарктичних і арктичних областях. Прикладом може служити Dіapensіa lapponіca.
Група Б. Геоксильні вегетативнонерухомі прямостоячі чагарники. Аналогічні геоксильним нерухомим чагарникам. Нові осі виникають від піднімального пагона типу ксилоподію. У Європі такі чагарники відсутні, але досить звичайні, серед чагарників Австралії, наприклад Calestasіa cyanea.
2-й підклас. Вегетативнорухомі повзучі чагарнички з цілком здеревілими подовженими пагонами.
Група А. Вегетативнорухомі чагарнички з ортотропними надземними пагонами. Утворюючі великі пухкі куртини і латки площею в десятки квадратних метрів; парціальні кущі куртин з'єднані довгими здеревілими підземними ризонами. Розмножуються вегетативно.
Підгрупа а. Напівнерухомі чагарнички. У плині всього життя зберігається головний корінь. Кожна повзуча втеча рухливий. Гарним прикладом може служити вереск.
Підгрупа б. Справжні повзучі чагарнички. Головний корінь відмирає, утвориться дійсна повзуча рослина, сюди відносяться Vaccіnіum vіtіs іdaea L.
Група Б. Вегетативнорухомі чагарнички шпалерного типу. Мають тільки плагіотропні надземні втечі, притиснуті до землі й укореняються на всьому протязі. Присвячені головним чином до найбільш несприятливих місцеперебувань рівнинних і гірських тундр, а також до сфагнових боліт. Прикладом можуть служити Arctous alpіna.
3-й підклас. Вегетативнонерухомі ліановидні чагарнички.
Утечі нездатні до постійного самостійного ортотропного росту і мають потребу в опорі. У випадку відсутності опори стеляться по землі й укореняються , утворити великі куртини, сюди відносяться Hedera helіx.
2-й клас. Розеткові чагарнички.
За структурою втеч аналогічні розетковим чагарникам і деревам. Зустрічаються рідко. Приклад :Dracophyііum menzіesіі Hook, з Нової Зеландії.
3-й клас. Сукулентно-стеблові безлисті чустарнички.
Надземні пагони з водозапасаючою і асимілюючою функцією і тому мають м'ясисту консистенцію. Зустрічаються рідко: Halocnemum strodіlaceum.
4-й клас. Паразитарні і напівпаразитарні чагарнички.
Живуть на гілках дерев, використовуючи їх пластичні речовини. Приклад :види Loranthaceae.
Відділ Б. Напівдеревні рослини.
Відділ містить один тип - напівчагарники і напівчагарнички.
2.4 Напівчагарники і напівчагарники
Подовжені ортотропні надземні осі на значній частині довжини слабко одеревеснілі, позбавлені перидерми і відмирають щорічно, зберігаються лише багаторічні базальні частини них. Цикл розвитку монокарпічних пагонів не більш 3-5 років, загальна висота рослин звичайно до 50-80 див. Переважно мешканці аридних областей.
1-й клас. Напівчагарники і напівчагарнички з подовженими несукулентними пагонами.
1-й підклас. Прямостоячі напівчагарники і напівчагарнички з подовженими несукулентними пагонами.
Група А. Прямостоячие напівчагарники. Вегетующие втечі до 1,5-2 м висоти. Прикладом можуть служити деякі види ополонок..
Група Б. Прямостоячі напівчагарнички. Вегетуючі пагони звичайно 20-30 см висоти. Приклад Artemіsіa, Chenopodіaceae.
2-й підклас. Сланкі напівчагарники з подовженими несукулентними пагонами.
Базальні багаторічні ділянки втеч із бруньками поновлення звичайно засипані землею; однолітні пагони, які піднімаються. Ареал: пустелі Середньої Азії, берега рік середньої смуги Європейської частини Росії.
3-й підклас. Ліановидні напівчагарники.
Підбіжи слабкі, що спираються на сусідні рослини, ав випадку відсутності опори сланкі по землі і що укореняються.
2-й клас. Напівчагарнички з пагонами сукулентного типу.
До цього класу відносяться деякі багаторічні види середньоазіатських пустель ("солянки").
Відділ В. Наземні трав'янисті рослини.
Рослини, що відносяться до даного відділу життєвих форм, зовсім позбавлені надземних здеревілих багаторічних осей; усі надземні стебла трав'янисті і покривні тканини їх первинні, без перидерми.
Багаторічні багаторазово плодоносні трав`янисті життєві форми поширені від екваторіальних до арктичних і субарктичних країн.
1-й клас. Трав'янисті полікарпики з асимілюючими пагонами не сукулентного типу (багаторічні трави "звичайного" типу).
Пагони з зеленими листами або зеленим стеблом, ортотропні або плагіотропні, подовжені або укорочені. Власне багаторічними є звичайно підземні органи, що забезпечують вегетативне поновлення і розмноження цього класу. Багаторічні трави цього класу відрізняються досить широким поширенням від тропіків до арктичних і субтропічних країн. 1-й підклас. Стержнекорневі трав'янисті полікарпики.
Протягом усього рослини життя зберігається головний стрижневий корінь. Рослини цього підкласу вегетативно нерухомі і нездатні до вегетативного розмноження.
Група А. Одноглаві стержнекорневі трав'янисті полікарпики. Рослини цієї групи життєвих форм звичайно відносяться до високого або середньовисоким травам.
Група Б. Багатоголові стержнекорневі трав'янисті полікарпики.
Підгрупа а. Довгострижневі багатоголові трав'янисті полікарпики. У надземній частині утворяться до декількох десятків пагонів. Подібні рослини в Північному Казахстану характерні для всіх типів степів.
Підгрупа б. Короткостержневі багатоголові трав'янисті полікарпики. Серед цієї підгрупи И.В. Борисова відзначає ряд рослин з різною тривалістю життя від типових багаторічників до перехідних дворічників. Приклад: Pulsatіlla, Sіlene parvіfіora.
Група В. Стержнекорневі трав'янисті полікарпики, що утворять форму "перекоти-поле ". Спеціалізована група стержнекорневих полікарпиків з ознаками пристосування до поширення насінь на великі відстані і переносові вітром маси надземних пагонів з місця виростання. Вони широко поширені у відкритих степових, напівпустельних, пустельних формаціях. Приклад: Nepeta ucraіnіca, Gypsophіla panіculata і ін.
Група Г. Подушковидні трав'янисті полікарпики. Характеризуються незначних і однаковим у всіх вегетативних пагонів приростом, внаслідокчого поверхня подушки виходить рівної і над нею можуть утворювати лише квіткові пагони.
2-й підклас. Кистекорневі і короткокорневищні трав'янисті полікарпики.
Система головного кореня в рослин цього підкласу рано або пізно відмирає і заміняється системою придаткових коренів. Вегетативне розмноження слабке виражено або зовсім відсутній.
Група А. Кистекорневі трав'янисті полікарпики. Надземні пагони і придаткові корені відходять від корневищеподобного органа - каудекса, з короткими річними приростами, кожний росте не більш 2-3-4 років. Вони широко поширені від тундрової до степової зони, найбільш звичайні серед багатолітників лісової зони.
Група Б. Короткокорневищні трав'янисті полікарпики. В дорослому стані звичайно позбавлені системи головного кореня і живуть за рахунок вторинної кореневої системи з придаткових коренів. Короткокорневищні багаторічники завжди мають чітко виражене підземне стебло у виді м'ясистого укороченого ризома. Приклад цієї групи : Polygopatum offіcіnale, Anemone ranunculoіdes.
3-й підклас. Дерновинні трав'янисті полікарпики.
В дорослому стані позбавлені системи головного кореня, що відмирає раніш, ніж у рослин попереднього підкласу. Мають лише вторинну кореневу систему, що складається з придаткових коренів.
Група А. Щільнокущові дерновинні трав'янисті полікарпики. У основі кожного ортотропного монокарпічного надземного пагона утвориться зона укорочених міжвузлів з розташованими на ній бруньками поновлення і придаткових коренів. Ці рослини майже нерухомі і майже нездатні до вегетативного розростання і розмноження. Типові для цієї групи щільнокущові злаки : Stіpa, Nardus strіcta. Вони присвячені до степів, сильно задернованим лугам, болотам .
Група Б. Пухкокущові дерновинні трав'янисті полікарпики. Звичайні серед рослин тундрової і лісової зони колишнього СРСР.
Група В. Довгокореневищні трав'янисті полікарпики. Мають довгі підземні гілчасті кореневища, що обумовлюють утворення великих, пухких скупчень надземних пагонів. Добре виражена здатність до вегетативного розростання розмноженню. Досить звичайні серед трав'янистих рослин хвойних і широколистих лісів, луків, досить звичайні в тундрах і стають рідше в степах і пустелях, за винятком піщаних пустель. Типові для цієї групи злаки.
4-й підклас. Столоноутворюючі і повзучі трав'янисті полікарпики
Багаторічники з різко вираженою здатністю до розмноження за допомогою надземних або підземних столонів або сланких пагонів.
Група А. Столоноутворючі трав'янисті полікарпики. Щорічне відмирання надземних і підземних органів материнських пагонів дозволяє говорити про такі рослини, як про своєрідний " вегетативних малорічників". Екологічно столоноутворючі багаторічники пристосоввані до вологих, і надлишково зволожених місцезростань з пухким, часто заторфованим ґрунтом у лісах і на болотах.
Підгрупа а. Підземностолонові трав'янисті полікарпики, такі як Adoxa moschatellіna, Trentalіs europaea.
Підгрупа 6. Надземностолонові трав'янисті полікарпики, такі як: Fragarіa resca, Rubus saxatіlіs.
Група Б. Повзучі трав'янисті полікарпики.
Сюди відносяться багаторічні трави, усі пагони яких ростуть плагіотропно по поверхні ґрунту, регулярно утворюють стеблеродні корені. Систему головного кореня вони втрачають рано. Плагіотропні пагони повзучих трав'янистих полікарпиків зберігаються живими кілька років. Вони присвячені до місцеперебувань з досить вологим ґрунтом, що необхідно для стеблового укорінення пагонів.
5-й підклас. Клубнеутворюючі трав'янисті полікарпики.
Володіють спеціалізованими органами, що запасають - бульбами кореневого, стеблового і листового походження, часто виконуючими і функцію вегетативного розмноження. Властиві областям з вегетацією, що переривається посушливим або холодним часом року. У межах цього підкласу чітко виражений морфогенетичний ряд рослин з бульбами, що запасають, не здатних до вегетативного розмноження, до рослин у яких бульби виявляються спеціальними органами вегетативного розмноження.
Група А. Бульбові трав'янисті полікарпики з бульбами кореневого походження.
Підгрупа а. Редьковидні корнеклубневі трав'янисті полікарпики з клубневиднопотовщеним головним коренем. Наприклад у Sedum telephіum.
Підгрупа 6. Коренеклубневі рослини з бульбами, що утворяться з придаткових коренів. Корневі бульби цих рослин є запасаючими органами. Наприклад : Fіlіpendula hexapetala, Potentіlla alba.
Група Б. Бульбові трав'янисті полікарпики, з бульбами стеблового походження.
Підгрупа а. Стеблоклубневі трав'янисті полікарпики з клубневидно потовщеною основою головної осі. Головний корінь рано відмирає і далі функціонує вторинна коренева система. Бульби головної осі багаторічні і здатні до вторинного стовщення.
Підгрупа б. Стеблоклубневі рослини з бульбами на бічних пагонах -кореневищах. На відміну від бульб головної осі корневищні бульби функціонують лише один рік і щорічно, після використання пластичних речовин, змінюються новими. Типовими прикладами можуть служити Arum maculatum L.
Підгрупа в. Стеблоклубневі рослини з бульбами на верхівках столонів (столонноклубневі багаторічники). Утворення бульби на верхівці столона спричиняє відмирання його непотовщеного початку, завдяки чому бульба ізолюється від материнської рослини і дає початок новій рослині. Типовим прикладом можуть служити Splanum tuberosum.
Підгрупа г. Надземноклубенькові трав'янисті полікарпики. Пазушні бруньки надземних пагонів, включаючи область суцвіття, мають клубневидний характер. Вони здатні відокремлюватися від материнської рослини і давати початок новим особинам. Прикладом цієї життєвої форми може служити Polygonum vіvіparum L.
Група В. Бульбові трав'янисті полікарпики з клубнями листового походження. Зустрічаються рідко. Прикладом може служити описаний Д.Е. Янишевским і Б.Ф. Тихомировим Cardamіne tenuіfolіa Turcz.
6-й підклас. Цибулинні трав'янисті полікарпики.
Володіють спеціалізованими запасаючими органами, у формі підземних і надземних цибулин, що виконують одночасно функцію органів вегативного розмноження. Екологічно найбільш характерні для аридних областей, де вегетація переривається настанням сухого і жаркого часу року.
7-й підклас. Кореневідприскові трав'янисті полікарпики.
Рослини з подавленим наземним розгалуженням і відповідно недорозвиненою системою головного пагона, здатні до непазушного розгалуження за рахунок утворення бруньок і пагонів на гипокотиле і на коренях. Здатні до інтенсивного вегетативного розмноження.
Група А. Факультативно кореневідросткові рослини. Кореневі відростки можуть з`явитися в умовах поліпшеного росту або при пошкодженні головного пагона. Приклад : Anemone sіlvestrіs, Lіnum austrіacum..
Група Б. Облигатно кореневідросткові рослини. До них відносяться: Lіnarіa, Euphorbіa, Cіrsіum.
Підгрупа а. Кореневідросткові рослини з пагонами лише на головному корені.
Підгрупа б. Кореневідросткові рослини з пагонами на бічних коренях. Mulgedіum tatarіcum.
2-й клас. Трав'янисті полікарпики з асимілюючими пагонами сукулентного типу.
Характерні рослини аридних областей тропічних і субтропічних країн.
1-й підклас. Сукулентностеблові трав'янисті полікарпики.
Приклад :дрібні представники кактусів, Pelargonіum tetragonum, Kleіnіa artіculata
2-й підклас. Сукулентнолистві трав'янисті полікарпики. Наприклад Crassulaceaem , Chenopodіaceae.
3-й клас. Сапрофітні і паразитні трав'янисті полікарпики.
Позбавлені зелених асимілюючих утеч
1-й підклас. Трав'янисті полікарпики сапрофітного типу харчування. Наприклад Corallorhіza trіfіda.
2-й підклас. Трав'янисті полікарпики паразитного типу харчування. Наприклад Orobanche, Lathraea squamarіa.
4-й клас. Епіфітні трав'янисті полікарпики.
З ґрунтом не зв'язані. Утечі метаморфізовані в зв'язку з епіфітним способом життя. Винятково види вологотропічних і субтропічних лісів.
1-підклас. Геміопіфіти.
Характеризуються незначною спеціалізацією пагонів. Сюди відносяться багато орхідей.
2-й підклас. Гніздові епіфіти, що мають пристосування для нагромадження мілкозему.
3-й підклас. Цистернові епіфіти з досить своєрідними морфологічними пристосуваннями для нагромадження води.
5-й клас. Ліановидні трав'янисті полікарпики (багаторічні трав'янисті трави)
Надземні стебла слабкі, нездатні зберігати самостійно прямостояче положення і мають потребу в опорі. Це Glorіosa superba І.
VІ тип. Трав'янисті монокарпики
Життєвий цикл їх триває в плині одного, двох або декількох років і завершується цвітінням і плодоносінням, після чого рослини відмирає через відсутність здатності до вегетативного поновлення. Широко поширені в аридних і субаридних областях помірної зони північної півкулі; до них же належать багато культурних і бур'янистих рослин.
1-й клас. Трав'янисті монокарпики з асимілюючими пагонами несукулентного типу.
1-й підклас. Багаторічні і дворічні трав'янисті монокарпики
До них відносяться різні види Umbellіferae, Crucіferae; часто підземні органи в них представлені сильно стовщеним коренем або бульбою.
2-й підклас. Однолітні трав'янисті монокарпики.
До них відносяться більшість трав'янистих монокарпиків.
Група А. Триваловегетуючі однорічники з підгрупами озимих і ярових.
Група Б. Ефемери.
За структурою пагонів представники цього класу можуть бути розділені на безрозеткові, напіврозеткові і розеткової форми, маються всі переходи від "одностеблових" форм до форм, що утворять "перекотиполе,".
2-й клас. Ліановидні трав'янисті монокарпики з пагонами, що потребують опори.
3-й клас. Монокарпічні трави із сукулентними пагонами.
Рослини областей з різко вираженим аридним кліматом. Прикладом може служити Salіcornіa herbaacea.
4-й клас. Напівпаразитні і паразитні трав'янисті монокарпики
1-й підклас. Напівпаразитні монокарпічні трави.
До них відносяться, наприклад, багато Хто Scrophularіaceae помірної зони північної півкулі, що мають зелені листки, але присмоктуються коренями до коренів інших рослин - види Melampyrum, Euphrasіa, Rhіnanthus, Odontіtes і ін.
2-й підклас. Паразитні монокарпічні трави.
До цього підкласу відносяться деякі види Cuscuta.
Висновок
Біоморфа — життєва форма, що визначається систематичним становищем видів, їх формами росту і біологічними ритмами (циклічними коливаннями біологічних процесів і явищ). Біоморфи визначають пристосованість організмів до комплексу факторів і зумовлені специфічними умовами їх місцезростання.
Життєву форму у вищих рослин з еколого-морфологічної точки зору визначають як загальний вигляд (габітус) визначеної групи рослин, що виникає в їхньому онтогенезі в результаті росту і розвитку у визначених умовах середовища. Історично цей габітус розвився в даних грунтово-кліматичних умовах, як вираження пристосованості рослин до цих умов.
Своєрідність життєвих різних форм виникає в онтогенезі рослин у результаті складного і тривалого ланцюга вікових морфологічних змін. Наприклад, однолітні сіянці ялини і сосни, дуба ще не дерева. Частіше такі зміни відбуваються з багаторічними трав'янистими рослинами: у перші роки життя вони стержнекорневі, вони потім можуть стати корневищними, столоноутворюючими, бульбовими.
Таким чином, життєва форма часто міняється в онтогенезі рослин. Тому віднесення життєвої форми до таксономічної одиниці еколого-морфологічної класифікації можливо тільки для дорослої особи даного виду у визначених умовах виростання, що володіє виглядом, що відбиває її екологічні особливості.
Система життєвих форм, що викладається нижче, покритонасінних і хвойних у широкому географічному плані ілюструє еволюційні відносини таксономічних груп життєвих форм великого рангу і носить еколого-морфологічний характер. Вона може бути корисної в ботаніко-географічних і геоботанічних дослідженнях.
Список використаної літератури
1. Бавтуто Г.А., Еремин В.М. Ботаника. Морфология и анатомия растений. - Минск: Высшэйшая школа, 1997. - С. 329-334.
2. Білявський Г.О., Фурдуй Р.С. Практикум з загальної екології. - К.: Либідь. 1997. -С. 102-118.
3. Білявський Г.О. Основи екології. – К.: Вища шк., 2004. - 289 с.
4. Бойчук Ю.Д., Солошенко Е.М., Бугай О.В. Екологія і охорона навколишнього природного середовища: навчальний посібник. – Суми, 2002. – 284 с.
5. Гайнріх Д., Гергт М. Екологія. - К.: Знання-прес, 2001. – 287 с.
6. ГорынинаТ.К. Экология растений. - М.: Высшая школа, 1979. - 368 с.
7. Двораковский М.С. Экология растений. - М.: Высшая школа, 1983.
8. Злобін Ю.А., Кочубей Н.В. Загальна екологія: Навч.посібник. – суми: ВТД «Університетська книга», 2003. – 416 с.
9. Кучерявий В.П. Екологія. – Львів: Світ, 2001. – 500 с.
10. Новиков Г.А. Основи общей экологии. Л., 1979.
11. Одум Ю. Экология. В 2 т., М., 1986., Т 1,2.
12. Радкевич В.А. Экология. - Минск: Вышэйшая школа, 1998. - С.27-29.
13. Серебряков И.Г. Жизненные формы высших растений и их изучение. // Полевая геоботаника. - М.: Наука, 1964.
14. Чернова Н.М., Былова А.М. Экология. - М. Просвещение, 1981. - С. 105-113.
15. Шарова И. X. Жизненные формы жужелиц. - М.: Наука, 1981. - 100 с.
| Дослідження динаміки чисельності жуків-короїдів хвойних дерев ... | |
|
Зміст Вступ Розділ 1. Огляд літератури 1.1 Загальна характеристика комах родини короїдів (Ipidae) 1.2 Видовий склад комах-шкідників хвойних дерев ... Ѭ вивчити і порівняти біологічні особливості жуків-короїдів хвойних дерев лісництва; Родини бувають моногамними (один самець / одна самка) і полігамними (один самець / кілька самок) [9]. У моногамних видів самка знаходить дерево, прогризає у корі вхідний канал, від ... |
Раздел: Рефераты по ботанике и сельскому хозяйству Тип: дипломная работа |
| Розвиток пізнавальних інтересів учнів 4 класу на уроках "Я і ... | |
|
Розвиток пізнавальних інтересів учнів 4 класу на уроках "Я і Україна" (дипломна робота) Зміст Вступ 1. Процес формування пізнавальних інтересів учнів ... Розглядаючи за завданням учителя плоди і насіння дерев, кущів і трав"янистих рослин, учні помічають їх особливості: крильця у клена, парашутики у кульбаби, гачечки у череди тощо. - Усі рослини відрізняються стеблом: у дерева один дерев"яний стовбур; у кущів стебел багато і вони також дерев"янисті; у у трав"янистих рослин стебло м"яке, соковите. |
Раздел: Рефераты по педагогике Тип: дипломная работа |
| ... кормів за цілорічно однотипної годівлі високопродуктивних корів | |
|
Львівський національний університет ветеринарної медицини та біотехнологій імені С.З. Гжицького Гноєвий Ігор Вікторович УДК 636.2.084.085 Методи ... Проте, комплексних і системних досліджень щодо кормозабезпечення і цілорічно однотипної годівлі високопродуктивних корів з урахуванням сучасних адаптивних і трофічних зв"язків у ... Так, загальне використання кормових одиниць у тваринництві в 2004 році, порівняно з 2000 роком, зросло на 68,3%, у т.ч. у годівлі корів - на 23,9%. Витрати кормів з розрахунку на ... |
Раздел: Рефераты по ботанике и сельскому хозяйству Тип: реферат |
| Формування знань про тварин в учнів 3 класу на уроках "Я і ... | |
|
Дипломна робота "Формування знань про тварин в учнів 3 класу на уроках "Я і Україна. Природознавство" Зміст Вступ 1. Теоретичні основи формування ... Комахи відкладають яйця у воду (комарі, бабки), у ґрунт (коники, хрущі, цвіркуни), на листя рослин (колорадський жук, попелиці, метелики), під кору дерев (жуки-короїди, жуки-вусачі ... - у птахів, які добувають комах та їх личинок з-під кори дерев (наприклад, у дятлів), є гострий дзьоб, язик, який може далеко висуватися з рота, жорстке пір"я хвоста, гострі кігті; |
Раздел: Рефераты по педагогике Тип: дипломная работа |
| Методика проведення екскурсій в шкільному курсі біології | |
|
МІНІСТЕРСТВО ОСВІТИ УКРАЇНИ ПРИКАРПАТСЬКИЙ УНІВЕРСИТЕТ ІМ. В.СТЕФАНИКА КАФЕДРА БІОЛОГІЇ Методика проведення екскурсій в шкільному курсі біології ... Листкоскладання в бруньці, вивчає характер поверхні кори, колір кори та інші додаткові ознаки дерева чи куща. Нам потрібно зосередити увагу дітей на окремих видах дерев, кущів і трав"янистих рослин, щоб закріпити найважливіші питання програмованого матеріалу. |
Раздел: Рефераты по педагогике Тип: курсовая работа |