Курсовая работа: Деятельность центральной нервной системы
1. Общий план строения центральной нервной системы
Нервная система – это совокупность специальных структур, объединяющая и координирующая деятельность всех органов и систем организма в постоянном взаимодействии с внешней средой.
К центральной нервной системе (ЦНС) относятся головной и спинной мозг, которые состоят из серого и белого вещества. Серое вещество спинного и головного мозга – это скопление нервных клеток вместе с ближайшими разветвлениями их отростков. Белое вещество – это нервные волокна, отростки нервных клеток, которые имеют миелиновую оболочку (она придает волокнам белый цвет).
Структурно-функциональной единицей нервной системы является нервная клетка – нейрон. Формы и размеры нейронов разных отделов нервной системы могут варьировать, но для них характерно наличие тела и отростков – одного длинного (аксона) и множества древовидных коротких (дендритов). Аксон проводит импульсы от тела нейрона к периферическим органам или к другим нервным клеткам. Функция дендритов – проведение импульсов к телу нейронов от периферических рецепторов и других нейронов. Передача нервного импульса от одного нейрона к другому происходит в местах их контактов- синапсах.
нервный система нейрон мозг
2. Нейрон как основной структурный и функциональный элемент нервной системы
Нервная ткань состоит из нервных клеток – нейронов и глиальных клеток (Шванновских клеток).
2.1 Строение, типы и развитие нейронов
Нейрон, или нервная клетка - это функциональная единица нервной системы. Нейроны восприимчивы к раздражению, т.е. способны возбуждаться и передавать электрические импульсы от рецепторов к эффекторам. По направлению передачи импульса различают:
1) афферентные нейроны (чувствительные, сенсорные нейроны) - проводят информацию об ощущении (импульс) от поверхности тела и внутренних органов в мозг;
2) эфферентные нейроны (двигательные нейроны) – проводят импульс («команды») от головного и спинного мозга ко всем рабочим органам;
3) вставочные нейроны (ассоциативные) – анализируют информацию и вырабатывают решения.
Вставочный нейрон передает возбуждение от афферентного (чувствительного) нейрона на эфферентные, лежит в пределах ЦНС. Тела эфферентных нейронов находятся в ЦНС или на периферии – в симпатических, парасимпатических узлах. Аксоны этих клеток продолжаются в виде нервных волокон к рабочим органам (произвольным – скелетным и непроизвольным – гладким мышцам, железам).
Каждый нейрон состоит из сомы (клетки диаметром от 3 до 100 мкм, содержащей ядро и другие клеточные органеллы, погруженные в цитоплазму) и отростков, обычно одного длинного отростка - аксона и нескольких коротких разветвленных отростков - дендритов. По дендритам импульсы следуют к телу клетки, по аксону - от тела клетки к другим нейронам, мышцам или железам. Благодаря отросткам нейроны контактируют друг с другом и образуют нейронные сети и круги, по которым циркулируют нервные импульсы.
Рис.1
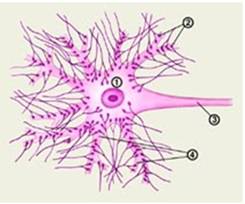
1 – тело (сома) нейрона; 2 – синаптические бляшки; 3 – аксон нейрона; 4 – дендриты.
На основании числа и расположения отростков нейроны делятся четыре группы:
1) униполярные нейроны – клетки с одним отрос тком - аксоном. Истинных униполярных клеток в теле человека нет, если не считать униполярной формы нейробластов до периода образования дендритов.
2) биполярные нейроны – клетки с двумя отростками - аксоном и дендритом. Истинные биполярные клетки в теле человека встречаются редко. Такую форму имеют часть клеток сетчатки глаза, спирального ганглия внутреннего уха и некоторые другие;
3) псевдоуниполярные нейроны – клетки, которые также имеют дендрит, идущий на периферию, заканчивающийся там чувствительными окончаниями (рецепторами), и аскон, несущий нервный импульс от тела клетки в ЦНС. Псевдоуниполярными они называются потому, что аксон и дендрит этих клеток начинаются от общего выроста тела, создающего впечатление одного отростка, с последующим Т-образным делением его.
4) мультиполярные (двигательные) нейроны. Из многих отростков такого нейрона один представлен аксоном, тогда как все остальные являются дендритами.
Нейрон развивается из небольшой клетки - предшественницы, которая перестает делиться еще до того, как выпустит свои отростки. Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма ее может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении - некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединенными друг с другом мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счет добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста - это область быстрого экзоцитоза и эндоцитоза, о чем свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путем экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
2.2 Глиальные клетки
Помимо нейронов нервная ткань содержит клетки еще одного типа. Они выполняют опорную и защитную функции и называются глиальными клетками или глией. По численности их в 10 раз больше, чем нейронов, и они занимают половину объема ЦНС. Глиальные клетки окружают нервные клетки и играют вспомогательную роль. Глиальные клетки более многочисленные, чем нейроны: составляют по крайней мере половину объема ЦНС ( рис. 1-18).
Глия не только выполняет опорные функции, но и обеспечивает многообразные метаболические процессы в нервной ткани, участвует в формировании миелиновой оболочки и способствует восстановлению нервной ткани после травм и инфекций.
Клетки нейроглии делятся на несколько типов. Клетки эпендимы выстилают желудочки головного мозга и спинномозговой канал и образуют эпителиальный слой в сосудистом сплетении. Они соединяют желудочки с нижележащими тканями.
Клетки макроглии делятся на две категории - астроциты и олигодендроциты .
Протоплазматические астроциты локализованы в сером веществе; от тела клетки, содержащей овальное ядро и большое количество гликогена, отходят сильно разветвленные короткие и толстые отростки.
Фибриллярные астроциты локализованы в белом веществе. Ядро у них также овальное, и тело клетки содержит много гликогена, но отростки длинные и менее разветвленные, некоторые ветви буквально упираются в стенки кровеносных сосудов. Эти клетки переносят питательные вещества из крови в нейроны.
Астроциты двух типов взаимосвязаны и образуют обширное трехмерное пространство, в которое погружены нейроны. Они часто делятся, образуя в случае повреждений центральной нервной ситемы рубцовую ткань.
Олигодендроциты локализованы в сером и белом веществе. Они мельче астроцитов и содержат одно сферическое ядро. От тела клетки отходит небольшое число тонких веточек, а само оно содержит цитоплазму с большим количеством рибосом. Шванновские клетки - это специализированные олигодендроциты, синтезирующие миелиновую оболочку миелинизированных волокон.
Клетки микроглии локализованы и в сером, и в белом веществе, но в сером веществе их больше. От каждого конца маленького продолговатого тела клетки, содержащей лизосомы и хорошо развитый аппарат Гольджи, отходит по толстому отростку. От всех его ветвей отходят более мелкие боковые веточки. При повреждении мозга эти клетки превращаются в фагоциты и, перемещаясь при помощи амебоидного движения, противостоят вторжению чужеродных частиц.
2.3 Взаимодействие глии и нейронов
Между нейронами и глиальными клетками существуют сообщающиеся между собой щели размером 15-20 нм, так называемое интерстициальное пространство, занимающее 12-14% общего объема мозга.
Во время потенциала действия концентрация ионов калия в интерстициальном пространстве может возрастать от 3-4 ммоль/л до 10 ммоль/л, что может вызвать значительную деполяризацию нервных клеток. В результате же активного транспорта ионов калия его внеклеточная концентрация может стать ниже нормальной, что вызывает гиперполяризацию нервных клеток.
Глиальные клетки имеют высокую проницаемость для ионов калия. Когда несколько глиальных клеток деполяризуются вследствие местного повышения концентрации ионов калия, между деполяризованными и недеполяризованными клетками возникает ток, создающий вход ионам калия в деполяризованные глиальные клетки, в результате чего внеклеточная концентрация ионов калия уменьшается. Благодаря высокой проницаемости глиальных клеток для ионов калия и электрическим связям между ними глиальные клетки действуют как буфер в случае повышения внеклеточной концентрации калия. Данных об активном поглощении ионов калия путем ионного насоса в глиальных клетках нет, хотя, возможно, они активно поглощают нейромедиаторы в некоторых синапсах, ограничивая таким образом время действия медиатора.
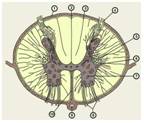
Рис.2
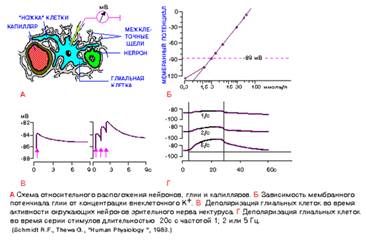
СВОЙСТВА ГЛИАЛЬНЫХ КЛЕТОК
А. Схема относительного расположения нейронов, глии и капилляров, составленная по электронно - микроскопическим данным. Глиальная клетка - астроцит (на рисунке желтая), в которую введен микроэлектрод для регистрации мембранного потенциала, находится между капилляром и нейроном. Все клеточные элементы разделены межклеточными промежутками шириной порядка 15 нм (на схеме относительная ширина щелей увеличена).
Б. Зависимость мембранного потенциала глии (ордината) от концентрации внеклеточного К+ . Средний уровень потенциала покоя составляет -89 мВ. Экспериментальные данные соответсвуют величинам потенциала, рассчитанным по уравнению Нернста, за исключением данных в области концентраций калия ниже 0,3 ммоль/л.
В. Деполяризация глиальных клеток во время активности окружающих нейронов зрительного нерва. Стимулы (с интервалом 1 с) показаны вертикальными стрелками.
Г. Деполяризация глиальных клеток в том же препарате во время серии стимулов длительностью 20 с с частотой 1, 2 и 5 Гц, в последнем случае деполяризация достигала почти 20 мВ. Следует обратить внимание, что на рис. В и Г временной ход деполяризации гораздо медленнее по сравнению с потенциалом действия.
2.4 Синапс
Синапс (греч. synapsis соприкосновение, соединение) — специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически синапс образован контактирующими мембранами двух клеток. Мембрана, принадлежащая отросткам нервных клеток, называется пресинаптической, мембрана клетки, к которой передается сигнал, — постсинаптической.
Схема строения межнейронного синапса:
Рис.3
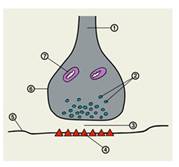
1 — нервное волокно (аксон); 2 — везикулы или синаптические пузырьки; 3 — синаптическая щель; 4 — рецепторы для медиатора; 5 — постсинаптическая мембрана; 6 —пресинаптическая мембрана; 7 — митохондрия.
В соответствии с принадлежностью постсинаптической мембраны синапсы подразделяют на нейросекреторные, нейромышечные и межнейроннные. Последние в зависимости от места их расположения разделяют на аксодендритические, аксосоматические, аксо-аксональные и дендро-дендритические. Различают аксодендритные синапсы (синапс между концевыми веточками аксона одного нейрона и дендритами другого нейрона) и аксосоматические синапсы (синапс между концевыми веточками аксона одного нейрона и телом другого нейрона).
Рис.4
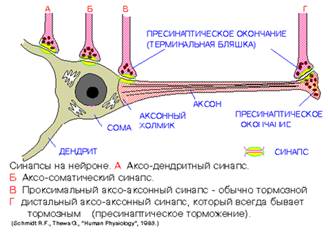
В зависимости от природы проходящих через синапсы сигналов, синапсы делятся на электрические синапсы (так называемые эфапсы) и химические синапсы. В синапсах с химической передачей возбуждения между пре- и постсинаптической мембранами имеется синаптическая щель, куда выделяется химическое вещество-передатчик — медиатор. Химические синапсы часто обозначают по названию медиатора (например, холинергические, адренергические, серотонинергические и т.п.). В эфапсе пре- и постсинаптические мембраны плотно соприкасаются и возбуждение передается посредством электрического тока. В зависимости от изменения биопотенциала постсинаптической мембраны различают синапсы деполяризующие, или возбуждающие, и гиперполяризующие, или тормозные.
Число синапсов очень велико, что обеспечивает большую площадь для передачи информации. На дендритах и телах отдельных двигательных нейронов спинного мозга находится свыше 1 000 синапсов. Некоторые клетки головного мозга могут иметь до 10 000 синапсов.
2.5 Механизм передачи возбуждения
Механизм передачи возбуждения принципиально одинаков во всех химических синапсах. В нем можно выделить следующие основные этапы: синтез и депонирование медиатора в пресинаптическом нейроне и его окончаниях; высвобождение медиатора из депонирующих везикул и его выход в синаптическую щель; взаимодействие медиатора со специфическими хеморецепторами постсинаптической мембраны с последующей генерацией биоэлектрического потенциала; инактивация выделенного медиатора с помощью ферментов или системы обратного поглощения. Биологически активные вещества, выполняющие функцию медиаторов, делят на несколько групп. К классическим нейромедиаторам относят ацетилхолин, адреналин и норадреналин, дофамин, серотонин, а также аминокислоты глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды. В соответствии с принципом Дейла, каждый отдельный нейрон в своих синаптических окончаниях один и тот же медиатор, поэтому нейроны можно обозначать также по виду медиатора: холинергические, адренергические, ГАМК-эргические, пептидергические. Синтез нейромедиаторов осуществляется как в соме нейрона с последующим аксонным транспортом, так и непосредственно в пресинаптических окончаниях аксона, где медиатор концентрируется в везикулах, или синаптических пузырьках.
В состоянии функционального покоя в пресинаптическом окончании происходит случайный контакт синаптических пузырьков с пресинаптической мембраной и выделение в синаптическую щель порции (кванта) медиатора из отдельной везикулы. Например, установлено, что в С. млекопитающих квант классического медиатора 10×ацетилхолина насчитывает 4210×—44 молекул. Выделившийся в синаптическую щель медиатор взаимодействует с хеморецепторами постсинаптической мембраны и приводит к возникновению миниатюрного постсинаптического потенциала. Приходящий к пресинаптическому окончанию потенциал действия в несколько раз увеличивает количество выделяемого в синаптическую щель медиатора. Взаимосвязь между потенциалом действия пресинаптической мембраны и процессом выделения медиатора из везикул в синаптическую щель обеспечивается ионами Са++. Выделяющийся в синаптическую щель медиатор взаимодействует с различными хеморецептивными участками на постсинаптической мембране. Роль мембранных, или клеточных, рецепторов играют белковые молекулы, обладающие способностью «узнавать» специфические для них вещества и вступать с ними в реакцию. В этом процессе большое значение придается системе аденилатциклаза-циклический АМФ. Предполагается, что взаимодействие медиатора с рецепторным участком аденилатциклазы приводит к образованию циклического АМФ с последующим повышением активности протеинкиназ цитоплазмы и ядра клетки. Вследствие этого интенсифицируется фосфорилирование белков, белковые молекулы подвергаются конформационным изменениям и происходит активация специальных ионных каналов мембраны. При увеличении проницаемости постсинаптической мембраны для ионов натрия, калия и хлора возникает ее деполяризация, регистрируется возбуждающий постсинаптический потенциал (ВПСП). При увеличении проницаемости лишь для ионов калия и хлора мембрана гиперполяризуется и регистрируется тормозный постсинаптический потенциал (ТПСП). ВПСП могут суммироваться, и при достижении величины деполяризации мембраны критического уровня генерируется потенциал действия. ТПСП тормозит генерацию потенциала действия, уменьшая суммарную величину ВПСП.
3. Строение и функции спинного мозга
Спинной мозг иннервирует скелетную мускулатуру (кроме мышц головы) и внутренние органы. Афферентными (чувствительными) путями спинной мозг связан с рецепторами, а эффернтными – со скелетной мускулатурой и со всеми внутренними органами. Основные функции спинного мозга:
1. рефлекторная (принимает участие в двигательных реакциях)
- здесь располагаются центры безусловных рефлексов (коленный рефлекс);
- вегетативные центры рефлексов мочеиспускания, дефекации, рефлекторная деятельность желудка.
2. проводниковая (проведение нервных импульсов)
- осуществляется связь различных отделов спинного мозга;
- связь головного мозга с остальными частями ЦНС;
- соединение рецепторов с исполнительными органами.
По внешнему виду спиной мозг представляет собой продолговатый, несколько плоский цилиндрический тяж длиной 45 см, диаметром 1 см . Он расположен в позвоночном канале и на уровне нижнего края большого затылочного отверстия переходит в головной мозг. В центре спинного мозга находится канал, заполненный спинномозговой жидкостью. Спинной мозг состоит из 31-32 сегментов: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1-2 копчиковых. Каждому сегменту спинного мозга соответствует отдельный участок тела, который иннервируется от спинномозгового нерва определенного сегмента.
Рис.5
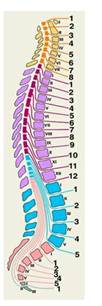
Схематическое изображение соотношений сегментов спинного мозга и позвонков на сагиттальном разрезе позвоночника. Оранжевым и желтым цветом обозначены шейные сегменты и шейные позвонки, фиолетовым и сиреневым — грудные, голубым — поясничные и копчиковые, розовым — крестцовые. Римскими цифрами обозначены позвонки, арабскими — корешки спинномозговых нервов соответствующих сегментов.
Вдоль всей передней поверхности спинного мозга в срединной сагиттальной плоскости тянется передняя срединная щель, а вдоль задней поверхности – задняя срединная борозда, которые разделяют спинной мозг на две симметричные половины. На его поверхности находятся две передние латеральные борозды, из которых выходят передние корешки, а на задней поверхности есть задние латеральные борозды – места входа с обеих сторон в спинной мозг задних корешков. Передние корешки заключают в себе только двигательные волокна, потому называются еще двигательными, задние - только чувствительные волокна, потому называются чувствительными корешками. Передние и задние корешки нервов проходят через межпозвоночные отверстия, где сливаются в один общий ствол, причем каждый задний корешок снабжается ганглиозным утолщением; нижние крестцовые пары выходят через передние и задние отверстия крестцовой кости. По выходе из позвоночника каждый нервный ствол разделяется на ветви, которые сперва имеют двигательные и чувствительные волокна и только при дальнейших разветвлениях дифференцируются, из 31-й пары нервов спинного мозга первые 8 пар называются шейными, следующие 12- грудными или спинными, 5 крестцовых и 1 копчиковая или хвостцовая. Между соседними нервами на некоторых местах образуются соединения посредством веточек, составляющие так наз. нервные сплетения. От этих сплетений и отходят в различные части тела главные нервные стволы, снабжающие эти части чувствительными или двигательными волокнами.
Серое вещество содержит нервные клетки и на поперечном разрезе напоминают форму бабочки. На протяжении всего спинного мозга серое вещество образует две вертикальные колонны, которые располагаются с двух сторон центрального канала. В каждой колонне различают передний и задний столбы (или рога). На уровне нижнего шейного, всех грудных и двух верхних поясничных сегментов спинного мозга в сером веществе выделяют боковой столб, который в других отделах спинного мозга отсутствует. Серое вещество задних рогов имеет неоднородную структуру. Основная масса нервных клеток заднего рога образует студенистое вещество и собственное ядро, а у основания заднего рога хорошо очерченное прослойкой белого вещества – грудное ядро, которое состоит из крупных нервных клеток.
Рис.6
Поперечный разрез спинного мозга: 1 — мягкая оболочка; 2 — дорсолатеральная (задняя) борозда; 3 — промежуточная дорсальная (задняя) борозда; 4 — дорсальный (задний) корешок; 5 — дорсальный (задний) рог; 6 — боковой рог; 7 — вентральный (передний) рог; 8 — вентральный (передний) корешок; 9 — передняя спинальная артерия; 10 — вентральная (передняя) срединная щель.
Клетки всех ядер задних рогов серого вещества – это, как правило, вставочные, промежуточные нейроны (интернейроны), отростки которых идут в белое вещество спинного мозга и далее к головному мозгу. Промежуточная зона, расположенная между передними и задними рогами, сбоку представлена боковым рогом. В последнем находятся центры симпатической части вегетативной нервной системы.
Белое вещество находится снаружи серого вещества. Борозды спинного мозга разделяют белое вещество на симметрично расположенные слева и справа три канатика: передний, боковой и задний. Белое вещество представлено отростками нервных клеток. Совокупность этих отростков в канатиках спинного мозга составляют три системы пучков – трактов (проводников):
1) короткие пучки ассоциативных волокон, которые связывают сегменты спинного мозга, расположенные на разных уровнях;
2) восходящие (чувствительные, афферентные) пучки, направляющиеся к центрам головного мозга или к мозжечку;
3) нисходящие (двигательные, эфферентные) пучки, идущие от головного мозга к клеткам передних рогов спинного мозга.
В белом веществе задних канатиков находятся восходящие пути, а в передних и боковых канатиках проходят восходящие и нисходящие системы волокон.
Передний канатик включает следующие проводящие пути:
1) передний корково-спинномозговой (пирамидный) путь. Этот путь передает импульсы двигательных реакций от коры большого полушария головного мозга к передним рогам спинного мозга;
2) передний спинно-таламический путь – обеспечивает проведение импульсов тактильной чувствительности;
3) преддверно-спинномозговой – берет начало от вестибулярных ядер VIII пары черепных нервов, расположенных в продолговатом мозге. По волокнам пути идут импульсы, поддерживающие равновесие и осуществляющие координацию движения.
Боковой канатик спинного мозга содержит следующие проводящие пути:
1) задний спинно-мозжечковый – несет проприоцептивные импульсы в мозжечок;
2) передний спинно-мозжечковый – идет в кору мозжечка;
3) латеральный спинно-таламический – проводит импульсы болевой и температурной чувствительности;
4) латеральный корково-спинномозговой (пирамидный) – проводит двигательные импульсы от коры большого мозга к спинному мозгу;
5) красноядерно-спинномозговой – проводит импульсы автоматического (подсознательного) управления движениями и поддерживает тонус скелетных мышц.
Задний канатик содержит пути сознательной проприоцептивной чувствительности (сознательное суставно-мышечное чувство), которые направляются в головной мозг и корковый конец двигательного анализатора, передают информацию о состоянии мышц тела, его частей в пространстве. На уровне шейных и верхних грудных сегментов спинного мозга задние канатики промежуточной бороздой делятся на два пучка – Тонкий пучок Голля и клиновидный пучок Бурдаха.
Рис.7
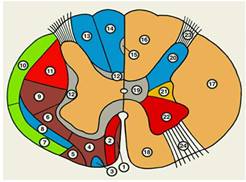
Схематическое изображение поперечного разреза спинного мозга.
Слева обозначены проводящие пути, справа — участки серого вещества; одинаковыми цветами обозначены группы проводящих путей и соответствующие им участки серого вещества; синим цветом — чувствительные пути и задний рог, красным — пирамидные пути и передний рог, серым — собственные пучки спинного мозга и промежуточное вещество, зеленым — восходящие пути экстрапирамидной системы, желтым — боковой рог: 1 — покрышечно-спинномозговой путь; 2 — передний корково-спинномозговой путь; 3 — передний спиноталамический путь; 4 — преддверно-спинномозговой путь; 5 — оливоспинномозговой путь; 6 — ретикул-спинномозговой путь: 7 — передний спиномозжечковый путь; 8 — латеральный спиноталамический путь; 9 — красноядерно-спинномозговой путь; 10 — задний спиномозжечковый путь; 11 — латеральный корково-спинномозговой путь; 12 — собственные пучки спинного мозга; 13 — клиновидный пучок; 14 — тонкий пучок; 15 — овальный пучок; 16 — задний канатик; 17 — боковой канатик; 18 — передний канатик: 19 — промежуточное вещество; 20 — задний рог; 21 — боковой рог; 22 — передний рог; 23 — задний корешок; 24 — передний корешок.
4. Головной мозг
Головной мозг, передний отдел центральной нервной системы позвоночных животных и человека, помещающийся в полости черепа. Головной мозг – материальный субстрат высшей нервной деятельности и главный регулятор всех жизненных функций организма. В целом головной мозг состоит из скоплений тел нервных клеток, нервных трактов и кровеносных сосудов. Нервные тракты образуют белое вещество мозга и состоят из пучков нервных волокон , проводящих импульсы к различным участкам серого вещества мозга - ядрам или центрам - или от них. Проводящие пути связывают между собой различные ядра, а так же головной мозг со спинным мозгом.
При осмотре препарата головного мозга хорошо заметны три его наиболее крупные составные части. Это парные полушария большого мозга, мозжечок и мозговой ствол.
В функциональном отношении мозг можно разделить на несколько отделов:
1) передний мозг, состоящий из конечного мозга (больших полушарий) и промежуточного мозга,
2)средний мозг, включающий ножки мозга и четверохолмие,
3)задний мозг, состоящий из мозжечка и варолиева моста ,
Продолговатый мозг, варолиев мост и средний мозг вместе называются стволом головного мозга.
4.1 Продолговатый мозг
Продолговатый мозг находится между задним и спинным мозгом. Нижняя граница продолговатого мозга соответствует уровню большого затылочного отверстия, или месту выхода корешков I пары спинномозговых нервов, верхняя граница проходит по заднему краю моста. Длина продолговатого мозга взрослого человека составляет в среднем 25 мм. Верхняя часть продолговатого мозга в отличие от нижней имеет некоторое утолщение, чем напоминает форму конуса. Борозды мозга являются продолжением борозд спинного мозга. По сторонам от передней срединной щели на вентральной поверхности продолговатого мозга расположены выпуклые, постепенно сужающиеся внизу пирамиды, образованные пирамидными трактами, часть волокон которых формирует перекрест пирамид. Латеральнее пирамиды с двух сторон находятся возвышения – оливы, отделенные от пирамиды передней латеральной бороздой, из которой выходят корешки подъязычного нерва (XII пара черепных нервов). В нижней части дорсальной поверхности продолговатого мозга проходит дорсальная срединная борозда, по сторонам которой заканчиваются утолщениями тонкий и клиновидный пучки задних канатиков спинного мозга, отделенные друг от друга задней промежуточной бороздой. В утолщениях пучков располагаются соответствующие ядра, от которых отходят волокна, формирующие медиальную петлю. Последняя на уровне продолговатого мозга образует перекрест. Пучки этого перекреста располагаются в межоливном слое, дорсальнее пирамид. Из заднелатеральной борозды продолговатого мозга выходят тонкие корешки языкоглоточного (IX пара), блуждающего (X пара) и добавочного (XI пара) черепных нервов, ядра которых лежат в дорсолатеральных отделах продолговатого мозга. На дорсальной поверхности части бокового канатика расширяются и вместе с волокнами от клиновидного и нижнего ядер образуют нижние мозжечковые ножки, ограничивающие снизу ромбовидную ямку; верхняя часть дорсальной поверхности участвует в образовании дна IV желудочка. Серое вещество продолговатого мозга представлено скоплениями нейронов, которые образуют нижние оливные ядра. Дорсальнее пирамид находится ретикулярная формация, состоящая из переплетений волокон – нервных клеток.
Рис.8
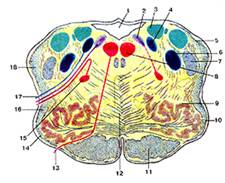
Продолговатый мозг. Поперечный разрез на уровне олив.
1-четвертый желудочек; 2-дорсальное ядро блуждающего нерва; 3-ядро вестибулярною нерва; 4-ядро одиночного пути; 5-задний (дорсальный) спинно-мозжечковый путь; 6-спинномозговое ядро тройничною нерва; 7-спинно-мозговой путь тройничного нерва; 8-ядро подъязычного нерва; 9-оливное ядро; 10-олива; 11-корково-спинномозговой путь (пирамидный); 12-медиальная петля; 13-подъязычный нерв; 14-передние наружные дуговые волокна; 15-двойное ядро; 16-спинно-таламический и спинно-покрышечный пути; 17-блуждающий нерв; 18-вентральпый (передний) спинно-мозжечковый путь.
Продолговатый мозг осуществляет рефлекторную и проводниковую функции. По чувствительным волокнам корешков черепных нервов он получает информацию (импульсы) от кожи, слизистых оболочек и органов головы, а также от рецепторов гортани, трахеи, внутренних органов грудной клетки (легкие, сердце), пищеварительной системы. Через продолговатый мозг осуществляются многие простые и сложные рефлексы. Например: 1) защитные – кашель, чиханье, рвота, слезоотделение, мигание; 2) пищевые – сосание, глотание, отделение пищеварительного сока; 3) сердечно-сосудистые, регулирующие деятельность сердца и кровеносных сосудов; 4) автоматически регулируемый дыхательный центр, обеспечивающий вентиляцию легких; 5) вестибулярные ядра, участвующие в осуществлении рефлексов позы, в перераспределении тонуса мышц.
Кроме того, через продолговатый мозг проходят пути, которые соединяют двусторонней связью кору головного мозга, промежуточный и средний мозг, мозжечок и спинной мозг.
4.2 Задний мозг
Задний мозг состоит из мозжечка и варолиева моста.
Мозжечок располагается кзади от моста и от верхней части продолговатого мозга, заполняя большую часть задней черепной ямки. В мозжечке различают верхнюю и нижнюю поверхности, границей между ними является задний край мозжечка, где проходит глубокая горизонтальная щель. Мозжечок лежит в дорсальной поверхности ствола мозга, охватывает его с боков и при помощи ножек соединяется с его частями: верхние мозжечковые ножки соединяют мозжечок со средним мозгом, средние- с мостом; нижние – с продолговатым мозгом. В мозжечке различают два полушария и непарную среднюю часть – червь мозжечка. На верхней и нижней поверхности полушарий и червя находится много параллельно идущих щелей мозжечка, между которыми находятся длинные и узкие извилины мозжечка.
Рис.9
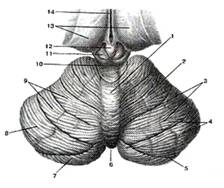
Мозжечок. Вид сверху. 1-червь мозжечка; 2-полушарие мозжечка; 3-щели (борозды) мозжечка; 4-листки мозжечка; 5-горизонтальная шель; 6-задняя вырезка мозжечка; 7-нижняя полулунная долька; 8-верхняя полулунная долька; 9-четырехугольная долька; 10-нижние холмики крыши среднего мозга; 11-верхний холмик; 12-эпифиз; 13-таламус; 14-третий желудочек.
Группы извилин, обособленные более глубокими бороздами, образуют дольки мозжечка. Полушария и червь мозжечка состоят из белого вещества, расположенного кнутри, и тонкой прослойки серого вещества коры мозжечка, которая охватывает белое вещество на периферии.
В коре мозжечка различают три слоя. Верхний слой называется молекулярным, так как на свежих срезах он испещрен мелкими точками. В нем расположены три типа нейронов - корзинчатые клетки, звездчатые клетки и клетки Лугаро. Направление аксонов клеток Лугаро неизвестно, аксоны корзинчатых клеток оканчиваются на теле, а звездчатых - на дендритах клеток Пуркинье .
Средний слой образован клетками Пуркинье, число которых у человека составляет 15 млн. Это крупные нейроны, их дендриты широко ветвятся в молекулярном слое. Аксоны клеток Пуркинье спускаются к ядрам мозжечка, и небольшое их количество заканчивается на вестибулярных ядрах. Это единственные аксоны, которые выходят из мозжечка.
Нижний слой коры мозжечка называется гранулярным, так как на срезах имеет зернистый вид. Этот слой составляют мелкие клетки-зерна (около 1 000-10 000 млн), аксоны которых идут в молекулярный слой. Там аксоны Т- образно делятся, посылая в каждом направлении вдоль поверхности коры ветвь ( параллельное волокно) длиной 1 -2 мм. Эти ветви проходят через области ветвления дендритов остальных типов нейронов мозжечка и образуют на них синапсы. В зернистом слое расположены также более крупные клетки Гольджи, дендриты которых распространяются на относительно далекие расстояния в молекулярном слое, а аксоны идут к клеткам-зернам.
В кору мозжечка входят два типа двигательных волокон. Лазящие (лиановидные) волокна проходят через зернистый слой и заканчиваются в молекулярном слое на дендритах клеток Пуркинье. Отростки лиановидных волокон оплетают дендриты этих клеток подобно ветвям плюща. К каждой клетке Пуркинье подходит только одно волокно, тогда как каждое лиановидное волокно иннервирует 10 - 15 нейронов Пуркинье. Все остальные афферентные пути мозжечка представлены гораздо более многочисленными (около 50 млн) мшистыми волокнами , оканчивающимися на клетках - зернах. Каждое мшистое волокно отдает множество коллатералей, благодаря чему одно такое волокно иннервирует множество клеток коры мозжечка. Вместе с тем к каждой клетке коры подходят многочисленные параллельные волокна от клеток-зерен, и поэтому через эти нейроны на любой клетке коры мозжечка конвергируют сотни мшистых волокон.
Основное значение мозжечка состоит в дополнении и коррекции деятельности остальных двигательных центров. Каждая из трех продольных зон мозжечка имеет свои функции. Червь мозжечка управляет позой, тонусом, поддерживающими движениями и равновесием тела. Промежуточный отдел мозжечка участвует во взаимной координации позных и целенаправленных движений и в коррекции выполняющихся движений. К полушариям мозжечка , в отличие от остальных его частей, сигналы поступают не непосредственно от периферических органов, а от ассоциативных зон коры головного мозга . Информация о замысле движения, передающаяся по афферентным путям к двигательным системам, превращается в полушариях мозжечка и его зубчатом ядре в программу движения, которая посылается к двигательным областям коры преимущественно через ядра таламуса . После этого становится возможным осуществление движения. Таким образом осуществляются очень быстрые движения, которыми невозможно управлять через соматосенсорные обратные связи.
Варолиев мост - вентральная часть заднего мозга, в нем проходят восходящие и нисходящие нервные пути. Кроме того, здесь имеются ядра, переключающие импульсы на мозжечок.
Мост снизу граничит с продолговатым мозгом, сверху проходит в ножки мозга, боковые его отделы образуют средние мозжечковые ножки. Из глубокой горизонтальной борозды, отделяющей мост от пирамид продолговатого мозга, выходят корешки отводящих нервов (VI пара), ядра которых лежат в дорсальном отделе моста. В латеральной части этой борозды видны корешки лицевого (VII пара) и преддверно-улиткового (VIII пара) нервов. Продолжение моста в латеральном направлении образует среднюю ножку моста. На вентральной поверхности моста имеется широкая, но неглубокая базилярная борозда, от нее с боков идут два продольных волокна, внутри которых проходят волокна пирамидных путей. Дорсальная поверхность моста прикрыта мозжечком и ее не видно снаружи. От дорсальной поверхности продолговатого мозга она отделяется мозговыми полосками и вместе с ней участвует в образовании ромбовидной ямки, или дна IV желудочка. Из переднебоковых отделов моста выходят пучки тройничного нерва, ядра которого лежат в дорсальной части моста и продолговатого мозга (V пара).
Рис.10
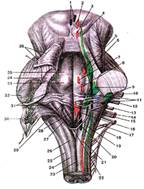
Ромбовидная ямка. Задняя поверхность моста и продолговатого мозга, проекция ядер (я)черепных нервов на ромбовидную ямку.
1-добавочное (парасимпатическое) я. глазодвигательного нерва; 2-я. глазодвигательного нерва; 3-я. блокового нерва; 4-я. среднемозгового пути тройничного норна; 5-двигательное я. тройничного нерва; 6-мостовое я. тройничного нерва; 7-я. отводящего нерва; 8-я. лицевого нерва; 9-я. преддверно-улиткового нерва; 10-корешок лицевого нерва (VII пара); 11-верхнее и нижнее слюноотделительные я.; 12-преддверно-улитковый нерв (VIII пара); 13-языкоглоточннй нерв (IX пара); 14-я. подъязычного нерва; 15-блуждащий нерв (X пара); 16-двойное ядро; 17-я. спинномозгового пути тройничного нерва; 18-добавочный нерв (XI пара); 19-ядро одиночного пути; 20-дорсальное ядро блуждающего нерва; 21-спинномозговое я. добавочного нерва; 22-задвижка; 23-задняя срединная борозда; 24-тонкий пучок; 25-клиновидный пучок; 26-бугорок топкою я.; 27-треугольник блуждающего нерва; 28-срединная борозда ромбовидной ямки; 29-мозговые полоски; 30-нижний мозговой парус (отвернут); 31-преддверное поле; 32-среднян ножка мозжечка; 33-лицевой бугорок; 34-верхняя ножка мозжечка; 35-срединное возвышение; 36-верхний мозговой парус (отвернут).
4.3 Средний мозг
Средний мозг связывает два передних отдела мозга с двумя задними отделами мозга, поэтому все нервные пути головного мозга проходят через эту область, составляющую часть ствола головного мозга. Крышу среднего мозга образует четверохолмие, где находятся центры зрительных рефлексов и слуховых рефлексов. Верхняя пара бугорков четверохолмия получает сенсорные импульсы от глаз и мышц головы и контролирует зрительные рефлексы. Нижняя пара бугорков четверохолмия получает импульсы от ушей и мышц головы и контролирует слуховые рефлексы.
На поперечном срезе среднего мозга хорошо выделяется своей темной окраской (за счет пигмента в клетках - меланина) черное вещество. Оно простирается в ножке мозга от моста до промежуточного мозга. В покрышке среднего мозга проходят восходящие проводящие пути и залегают ядра среднего мозга. Самым крупным ядром покрышки на разрезе среднего мозга является красное ядро. Оно находится несколько выше черного вещества, имеет продолговатую форму и простирается от уровня нижних холмиков до гипоталамуса.
Красное ядро – одно из центральных координационных образований экстрапирамидной системы. Основная функция этого ядра – управление мышцами-разгибателями (при повреждении наблюдается ригидность мышц).
Водопровод среднего мозга (сильвиев водопровод) – узкий канал длинной 1,5 см; соединяет полость III желудочка с IV и содержит спинномозговую жидкость. Вокруг водопровода среднего мозга находится центральное серое вещество, в котором расположены ядра III и IV пар черепных нервов.
Функциональное значение среднего мозга заключается в том, что здесь находятся подкорковые центры слуха и зрения; ядра черепных нервов, обеспечивающие иннервацию поперечнополосатых и гладких мышц глазного яблока; ядра, относящиеся к экстрапирамидной системе (черное вещество, красное ядро), которые обеспечивают сокращение мышц тела во время автоматических движений. Кроме того, через средний мозг проходят нисходящие (двигательные) и восходящие (чувствительные) проводящие пути. Область среднего мозга является также местом расположения вегетативных центров и ретикулярной формации.
4.4 Передний мозг
Передний мозг включает мозг конечный, промежуточный мозг и базальные ганглии.
Промежуточный мозг - это задний отдел переднего мозга, он состоит из:
1)таламуса,
2) гипоталамуса,
В таламусе оканчиваются аксоны большинства сенсорных нейронов, несущих импульсы в кору головного мозга. Здесь анализируется характер и происхождение этих импульсов, и они передаются в соответствующие сенсорные зоны коры по волокнам, берущим начало в таламусе. Таким образом, таламус играет роль перерабатывающего, интегрирующего и переключающего центра для всей сенсорной информации. Кроме того, в таламусе модифицируется информация, поступающая из определенных зон коры, и полагают, что он участвует в ощущении боли и ощущении удовольствия. В таламусе начинается та область ретикулярной формации, которая имееет отношение к регуляции двигательной активности. Дорсальный участок, лежащий непосредственно перед таламусом - переднее сосудистое сплетение - ответственен за транспорт веществ между спинномозговой жидкостью, находящейся в третьем желудочке, и жидкостью, заполняющей подпаутинное пространство.
Таламус можно разделить на несколько функционально и/или анатомически различных ядер. Большая часть таламических ядер связана массивными трактами с областями коры головного мозга. Ядра таламуса можно разделить на четыре класса (см. рис. ниже) (упрощенно):
1. Специфические ядра таламуса
2. Неспецифические ядра таламуса
3. Ядра таламуса с моторными функциями
4. Ядра таламуса с ассоциатиавными функциями
Специфическим соматосенсорным ядром является вентробазальное ядро; оно соединено с обеими соматосенсорными областями коры головного мозга.
Рис.11
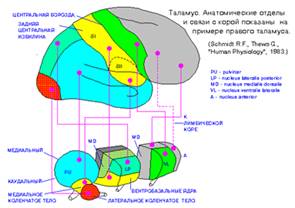
В связи с тем, что в здесь происходит переключение большей части чувствительных проводящих путей, таламус фактически является подкорковым чувствительным центром, а его подушка – подкорковым зрительным центром.
Гипоталамус расположен в основании переднего мозга непосредственно под таламусом и над гипофизом. Его вес составляет примерно 5 г. Гипоталамус не имеет четких границ, его можно рассматривать как часть сети нейронов, протягивающейся от среднего мозга через гипоталамус к глубинным отделам переднего мозга.
Гипоталамус - главный координирующий и регулирующий центр вегетативной нервной системы. К нему подходят волокна сенсорных нейронов от всех висцеральных рецепторов, вкусовых рецепторов и обонятельных рецепторов. Отсюда через продолговатый мозг и спинной мозг происходит регуляция сердечного ритма, регуляция кровяного давления, регуляция дыхания и регуляция перистальтики. В других участках гипоталамуса лежат специальные центры, от которых зависят голод, жажда и сон, а также поведенческие реакции, связанные с агрессивностью и поведенческие реакции, связанные с размножением. Гипоталамус контролирует концентрацию метаболитов и температуру крови, вместе с гипофизом регулирует секрецию большинства гормонов и поддерживает постоянство состава крови и постоянство состава тканей .
В лимбической системе и гипоталамусе, отвечающих за мотивации и эмоции, обнаружены рецепторы эстрадиола и тестостерона. Нервные клетки ядер гипоталамуса обладают способностью вырабатывать нейрогормоны (вазопрессин, или антидиуретический гормон, окситоцин), которые затем по разветвлениям аксонов нейросекреторных клеток поступают в заднюю долю гипофиза и током крови разносятся по организму. Некоторые ядра гипоталамуса вырабатывают так называемые рилизинг-факторы (либерины) и ингибирующие факторы (статины), регулирующие деятельность аденогипофиза. Последний передает информацию дальше в виде тропных гормонов периферическим железам внутренней секреции. Релизинг-фактор способствует высвобождению тирео-, лютео-, кортикотропина, пролактина, сомато- и меланотропина. Статины тормозят выделение последних двух гормонов и пролактина. Из гипоталамуса выделены также пептидовидные вещества энкефалины и эндорфины, которые обладают морфиноподобным действием.
Эпифиз, или верхний мозговой придаток, или шишковидная железа - эндокринный орган, расположенный между передними буграми четверохолмия над третьим мозговым желудочком. В эпифизе образуется гормон мелатонин, регулирующий пигментный обмен организма и обладающий антигонадотропным действием. Кроме мелатонина в пигментный обмен организма и обладающий антигонадотропным действием. Кроме мелатонина в эпифизе образуется и аккумулируется другое биологически активное, но негормональное производное триптофана - серотонин ( 5-оксиртриптамин), близкое по структуре к мелатонину и один из ближайших его предшественников в процессе биосинтеза. Серотонин отвечает за запуск сна.
Рис.12
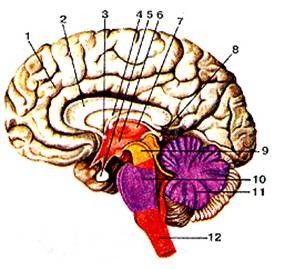
Головной мозг. Сагиттальный разрез. Вид с медиальной стороны.1-полушарие большого мозга; 2-мозолистое тело; 3-передняя (белая) спайка; 4-свод мозга; 5-гипофиз; 6-полость промежуточного мозга (III желудочек); 7-таламус; 8-эпифиз мозга; 9-средний мозг; 10-мост; 11 (-мозжечок; 12-продолговатый мозг.
4.5 Конечный мозг
Конечный мозг - это самый передний участок головного мозга, он состоит из двух полушарий большого мозга и базальных ганглий.
Большой мозг составляет крышу и стенки конечного мозга и достигает крупных размеров, образуя левое полушарие головного мозга и правое полушарие головного мозга, покрывающие сверху большую часть головного мозга.
Полушария большого мозга состоят из коры головного мозга и лежащей под ней центральной массы белого вещества головного мозга. Белое вещество состоит из проводящих путей. Левое и правое полушария соединены широким нервным трактом, который называется мозолистым телом. Каждое полушарие для удобства делят на четыре доли.
Кора головного мозга представляет собой тонкий слой нервной ткани, образующей множество складок. Общая поверхность коры составляет примерно 2200 кв.см. Толщина коры в различных частях больших полушарий колеблется от 1,3 до 4,5 мм, а общий объем составляет 600 куб.см. В состав коры входит 10 000 - 100 000 млн нейронов и еще большее число глиальных клеток (точное число которых еще не известно). В коре наблюдается чередование слоев, содержащих преимущественно тела нервных клеток, со слоями, образованными в основном их аксонами. Более 90% всех областей коры имеет типичное шестислойное строение и называется изокортексом . Слои нумеруются с поверхности вглубь:
1. Молекулярный слой коры головного мозга - образован волокнами, сплетенными между собой, содержит мало клеток.
2. Наружный зернистый слой коры головного мозга - характеризуется густым расположением мелких нейронов самой различной формы. В глубине располагаются малые пирамидные клетки (названные так благодаря своей форме).
3. Наружный пирамидный слой коры головного мозга - состоит в основном из пирамидных нейронов разной величины, более крупные клетки лежат более глубоко.
4. Внутренний зернистый слой коры головного мозга - характеризуется рыхлым расположением мелких нейронов различной величины, мимо которых проходят плотные пучки волокон перпендикулярно к поверхности коры.
5. Внутренний пирамидный слой коры головного мозга - состоит в основном из средних и больших пирамидных нейронов, апикальные дендриты которых простираются до молекулярного слоя.
6. Слой веретеновидных клеток коры головного мозга ( фузиформных клеток коры головного мозга) - в нем расположены веретеновидные нейроны, глубинная часть этого слоя переходит в белое вещество головного мозга.
На основании плотности, расположения и формы нейронов кора головного мозга делится на несколько полей, которые в некоторой степени совпадают с зонами, которым на основании физиологических и клинических данных приписывают определенные функции.
С помощью электрофизиологических методов установлено, что в коре можно различить области трех типов в соответствии с функциями, которые выполняют находящиеся в них клетки: сенсорные зоны коры головного мозга, ассоциативные зоны коры головного мозга и двигательные зоны коры головного мозга. Взаимосвязи между этими зонами позволяют коре большого мозга контролировать и координировать все произвольные и некоторые непроизвольные формы деятельности, включая такие высшие функции, как память, учение, сознание и свойства личности.
Сенсорные зоны через восходящие нервные пути получают сенсорную информацию от большинства рецепторов тела. Они занимают отдельные участки коры, связанные с определенными видами ощущений. Размеры этих зон коррелируют с числом рецепторов в соответствующей сенсорной системе.
- первичные сенсорные зоны и первичные моторные зоны (проекционные зоны);
- вторичные сенсорные зоны и вторичные моторные зоны (ассоциативные одномодальные зоны);
- третичные зоны (ассоциативные разномодальные зоны).
Первичные сенсорные и моторные зоны занимают менее 10% поверхности коры головного мозга и обеспечивают наиболее простые сенсорные и двигательные функции.
Ассоциативные зоны - это функциональные зоны коры головного мозга. Они связывают вновь поступающую сенсорную информацию с полученной ранее и хранящейся в блоках памяти, а также сравнивают между собой информацию, получаемую от разных рецепторов. Сенсорные сигналы интерпретируются, осмысливаются и при необходимости используются для определения наиболее подходящих ответных реакций, которые выбираются в ассоциативной зоне и передаются в связанную с ней двигательную зону. Таким образом, ассоциативные зоны участвуют в процессах запоминания, учения и мышления, и результаты их деятельности составляют то, что обычно называют интеллектом.
Отдельные крупные ассоциативные области расположены в коре рядом с соответствующими сенсорными зонами. Например, зрительная ассоциативная зона расположена в затылочной зоне непосредственно впереди сенсорной зрительной зоны и осуществляет описанные выше ассоциативные функции, связанные со зрительными ощущениями. Некоторые ассоциативные зоны выполняют лишь ограниченную специализированную функцию и связаны с другими ассоциативными центрами, способными подвергать информацию дальнейшей обработке. Например, звуковая ассоциативная зона анализирует звуки, разделяя их на категории, а затем передает сигналы в более специализированные зоны, такие как речевая ассоциативная зона, где воспринимается смысл услышанных слов.
Эти зоны относятся к ассоциативной коре и участвуют в организации когнитивных функций и сложных форм поведения.
Двигательные зоны - это е зоны коры головного мозга, посылающие двигательные импульсы к произвольным мышцам по нисходящим путям, которые начинаются в белом веществе больших полушарий.
Многие двигательные импульсы идут прямо в спинной мозг через два больших пирамидных тракта ( кортикоспинальных тракта ), проходящих в стволе мозга. Остальные двигательные импульсы передаются по экстрапирамидным путям, здесь же идут двигательные импульсы от базальных ганглиев и мозжечка . В продолговатом мозге все пути перекрещиваются, так что импульсы, идущие от коры левого полушария , иннервируют правую половину тела и наоборот.
Тела нейронов, участвующих в образовании пирамидных трактов, лежат в двигательных зонах коры, а их аксоны образуют синапсы непосредственно с мотонейронами спинного мозга в том его сегменте, где эти нейроны выходят на периферию. В головном мозге нет никаких промежуточных синапсов, поэтому импульсы и последующие ответы на них по пути не задерживаются и не видоизменяются.
Главным экстрапирамидным трактом является ретикулоспинальный тракт, переключающий импульсы от ретикулярной формации, которая лежит в стволе мозга между таламусом и продолговатым мозгом. Из различных отделов головного мозга, контролирующих двигательную активность, импульсы поступают в определенные участки ретикулярной формации, где они модифицируются под воздействием импульсов, идущих от коры, и становятся либо возбуждающими, либо тормозными. Например, импульсы от мозжечка и премоторной зоны коры, управляющей координированными движениями, поступают в ту область ретикулярной формации, которая находится в продолговатом мозгу и посылает импульсы, стимулирующие тормозные мотонейроны. Последние подавляют активацию определенных мышц, что дает возможность осуществлять сложные координированные движения тела. Другие комбинации двигательных импульсов, напротив, стимулируют возбуждающие нейроны, и общее воздействие ретикулярной формации на двигательную активность оказывается возбуждающим.
Большинство волокон сенсорных нейронов на своем пути через таламус к коре отдает коллатерали (боковые ветви) в ретикулярную формацию, участвуя в образовании ретикулярной активирующей системы, которая тонизирует кору и участвует в пробуждении организма от сна. Недостаточная активность этой системы или ее разрушение приводит соответственно к глубокому сну или коме . Как полагают, многие вещества, вызывающие общий наркоз, оказывают свое действие, временно блокируя синаптическую передачу в этой системе. Предполагается так же, что ретикулярная активирующая система ответственна за возникновение и поддержание побуждений к действию и концентрации внимания.
Функции некоторых участков коры, в частности обширных передних областей - префронтальных зон коры головного мозга - остаются еще неясными. Эти области, а также ряд других участков мозга, называют немыми зонами коры головного мозга, так как при раздражении их электрическим током не возникает никаких ощущений или реакций. Предполагают, что эти зоны ответственны за наши индивидуальные особенности, или личность. Удаление этих зон или перезку проводящих путей, идущих от них к остальному мозгу (префронтальную лоботомию) применяли для снятия у больных острого возбуждения, но от этого пришлось отказаться из-за таких побочных эффектов, как снижение уровня сознания и интеллекта, способности к логическому мышлениию и способности к творчеству. Эти побочные эффекты косвенно указывают на функции, выполняемые префронтальными зонами.
Базальные ганглии расположены в основании конечного мозга и представляют собой важное подкорковое связующее звено между ассоциативными областями коры головного мозга и двигательными областями коры головного мозга. К базальным ганглиям относятся следующие структуры: полосатое ядро базальных ганглий (стриатум), состоящее из хвостатого ядра базальных ганглий и скорлупы базальных ганглий, бледный шар базальных ганглий (паллидум), подразделяющийся на внутренний и внешний отделы, черная субстанция базальных ганглий и субталамическое ядро базальных ганглий. В состав базальных ганглиев часто включают также ограду и реже - миндалину.
Рис.13

Базальные (подкорковые) узлы и внутренняя капсула на горизонтальном разрезеголовного мозга. Вид сверху.
1-кора большого мозга (плащ); 2-колено мозолистого тела; 3-пе-редний рог бокового желудочка; 4-внутренняя капсула; 5-каруж-ная капсула; 6-ограда; 7-самая наружная капсула; 8-скорлупа; 9-бледный шар; 10-III желудочек; 11-задний рог бокового желудочка; 12-thalamus (зрительный бугор); 13-корковое вещество (кора) островка; 14-головка хвостатого ядра; 15-полость прозрачной перегородки.
Базальные ядра облегчают запускаемые корой движения и подавляют лишние сопутствующие движения. Стриатум получает соматотопически организованные проекции почти от всех зон коры. Эти проекции организованы в виде параллельных путей, которые начинаются от лобных областей, постцентральных (соматосенсорных) областей, прецентральных (моторных) областей и теменно-височно-затылочных областей. В то же время прямое отношение к регуляции движений имеют только глутаматергические, возбуждающие проекции от моторной коры и соматосенсорной коры .
Нейроны стриатума посылают волокна к структурам, формирующим главный эфферентный путь базальных ядер - к сетчатой части черной субстанции и медиальному бледному шару. Эти структуры, в свою очередь, образуют тормозные ГАМК ергические синапсы на переднем и латеральном вентральных ядрах таламуса, которые направляют к коре возбуждающие глутаматергические волокна.
Таким образом, при торможении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору усиливается - что и способствует облегчению запускаемого корой движения.
Напротив, при возбуждении нейронов сетчатой части черной субстанции и медиального бледного шара возбуждающее влияние таламуса на кору подавляется - что приводит к торможению ненужного движения.
Прямого выхода на спинной мозг базальные ядра не имеют. Прямой путь от стриатума к сетчатой части черной субстанции и медиальному бледному шару образован тормозными ГАМКергическими волокнами. Функция их состоит в усилении возбуждающего влияния ядер таламуса на те отделы моторной коры, которые отвечают за нужное движение.
Непрямой путь организован сложнее. Функция его заключается в подавлении возбуждающего влияния таламуса на другие отделы моторной коры. Первое звено этого пути - тормозные ГАМКергические проекции стриатума на латеральный бледный шар. Латеральный бледный шар посылает тормозные ГАМКергические волокна к субталамическому ядру. Выходы субталамического ядра представлены возбуждающими глутаматергическими волокнами - часть их возвращается к латеральному бледному шару, другие направляются к сетчатой части черной субстанции и медиальному бледному шару.
Таким образом, если активация прямого пути от стриатума усиливает возбуждение моторной коры, то активация непрямого пути - ослабляет.
Активность этих двух путей регулирует компактная часть черной субстанции, посылающая к стриатуму дофаминергические волокна. Эти волокна возбуждают прямой эфферентный путь стриатума (через нейроны с D1-рецепторами) и тормозят - непрямой (через нейроны с D2-рецепторами). Стриатум посылает также тормозные ГАМКергические волокна к компактной части черной субстанции, образуя тем самым с ней отрицательную обратную связь. Наконец, активность стриатума модулируют его холинергические вставочные нейроны - антагонисты дофаминергических нейронов.
4.6 Лимбическая система
Лимбическая система. Второй уровень центральной переработки информации происходит в четырех функциональных системах, к которым приходят сигналы от органов чувств после предварительной переработки в центральной нервной системе. Это ассоциативная система, лимбическая система, двигательная система и вегетативная система. Сложное, почти неисследованное взаимодействие этих областей мозга можно считать основой нашего поведения. Лимбические и паралимбические структуры,передние и медиальные ядра таламуса, медиальные и базальные отделы стриатума, а также гипоталамус формируют обширную нейронную структуру - лимбическую систему. Она координирует эмоциональные, мотивационные, вегетативные и эндокринные процессы.
Лимбическая система обеспечивает и еще одну важную функцию, нарушение которой часто встречается в клинической практике, - декларативную, или вербальную, память о событиях, приобретенных навыках и накопленных знаниях.
Несмотря на то, что поражение лимбических структур вызывает амнезию, лимбическую систему нельзя считать хранилищем. Следы памяти распределены по всей ассоциативной коре, и роль лимбической системы состоит в объединении этих отдельных фрагментов в доступные для припоминания события и знания. Поражение лимбической системы не стирает следы памяти, а нарушает их сознательное воспроизведение, при этом отдельные фрагменты информации остаются сохранными и обеспечивают так называемую процедурную память. Так, больные с корсаковским синдромом способны усвоить новый двигательный или перцептивный навык, но при этом не помнят, как и чему научились
| Неотложная помощь в невропатологии | |
|
Неотложная помощь в невропатологии Часть первая НЕВРОЛОГИЧЕСКИЕ СИНДРОМЫ ГОЛОВНАЯ БОЛЬ Механизмы возникновения. Многие внутричерепные образования ... Параличи и парезы возникают при поражениях периферического двигательного нейрона (передние рога спинного мозга, передние корешки, периферические нервы), характеризуются мышечной ... При полиомиелите поражаются преимущественно мотонейроны передних рогов спинного мозга и ядер черепных нервов мозгового ствола, а также нервные клетки коры больших полушарий ... |
Раздел: Рефераты по медицине Тип: книга |
| Оружие геноцида: самоубийство людей и его механизмы | |
|
Академия управления глобальными и региональными процессами социального и экономического развития ПРОГНОЗНО-АНАЛИТИЧЕСКИЙ ЦЕНТР Оружие геноцида ... В случаях острого алкогольного отравления, но не смертельного, в головном мозгу и в нервных клетках коры бывают те же изменения, что описаны выше, приводящие к глубоким изменениям ... 16] Нейрон (от греческого neuron - нерв) - нервная клетка, основная структурная и функциональная единица нервной системы; состоит из тела и отходящих от него отростков ... |
Раздел: Рефераты по медицине Тип: книга |
| Физиология высшей нервной деятельности | |
|
Т.Г. Анищенко О.В. Глушковская-Семячкина Л.Н. Шорина Н.Б. Игошева ФИЗИОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Т.Г. Анищенко, О.В. Глушковская-Семячкина, Л ... С позиций рефлекторного принципа были исследованы функции спинного мозга и различных отделов головного мозга - продолговатого, среднего, промежуточного мозга, мозжечка. Информация от кожных рецепторов, проприорецепторов и интерорецепторов идет к головному мозгу по нервным волокнам мощных восходящих путей в составе задних и частично боковых ... |
Раздел: Рефераты по биологии Тип: реферат |
| Сборник экзаменационных билетов по медицинским предметам и психологии | |
|
Экзаменационный билет по предмету АНАТОМИЯ ЦНС Билет № 1 Особенности формирования таламуса и глазного яблока. Фиброзные астроциты и их расположение ... Строение передних и задних корешков спинного мозга. Классификация нейронов в коре головного мозга. |
Раздел: Рефераты по медицине Тип: реферат |
| Шпоры по анатомии и физиологии человека | |
|
ПРЕДМЕТ И СОДЕРЖАНИЕ ВОЗРАСТНОЙ ФИЗИОЛОГИИ Термин "ГИГИЕНА" происходит от греческого слова - целебный, *приносящий здоровье. Его происхождение связано ... собой нейронов^ Она представлена головным и спинным мозгомД На разрезе головного и спинного мозга различают участки более темного цвета - серое вещество (образовано телами нервных ... Миелизация раньше всего отмечена у периферических нервов, затем ей подвергаются волокна опийного мозга, стволовой части головного мозга, мозжечка и позже волокна больших -полушарий ... |
Раздел: Рефераты по биологии Тип: реферат |